

15. Динамични свойства на мускулите. Механизъм на мускулното съкращение.
Мускулното съкращение е най-добре изученото механо-химично явление. Гръбначните животни имат два вида основни вида мускули – гладки и напречно-набраздени. Те се различават по форма и по големина и притежават функционални и морфологични различия, но съкратителният апарат в тях е еднакъв по строеж и функции.
Отделното мускулно влакно има диаметър 0.05-0.1mm и съдържа около 1000-2000 съединителни влакна, всяко с диаметър 1-2μm, наречени миофибрили. Миофиб-рилите от своя страна са изградени от надлъжно ориентирани белтъчни актинови и миозинови молекули (нишки). Не при всички мускули нишките са обединени в миофибрили. В някои те са разположени в цитоплазмата на мускулната клетка.
При наблюдение на миофибрили от напречнонабраздени мускули с поляризационен микроскоп се вижда напречно набраздяване – светли и тъмни участъци. Тъмните са анизотропни двойнопречупваци и се наричат А-зона. Изградени са от белтъка миозин. Светлите ивици са изотропни, изградени основно от белтъка актин и се наричат I-зона. В средата на всяка I-зона има тъмен Z-диск, представляващ тънка мембрана, през чиито пори преминават миофибрилите. Участъкът, ограничен от две Z диска, се нарича саркомер. В центъра на А-зоната е разположена хиалинна зона (Н-зона) – свободно от актинови нишки междинно пространство (Фиг. 15.1).
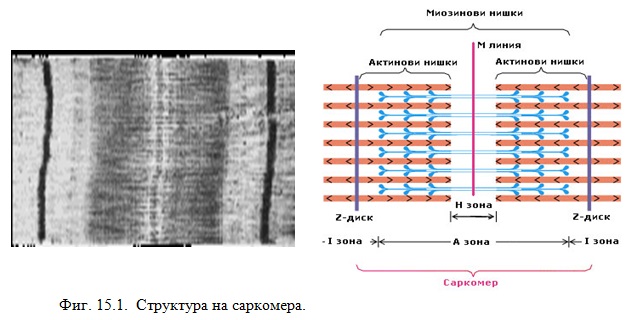
Описаната структура се наблюдава в напречнонабраздените мускули на гръбначните животни, които са специализирани за бързи и краткотрайни съкращения. Освен тях има и мускули, при които под поляри-зационен мисроскоп не се наблюдава напречно набраздяване. Тяхното съкращаване се извършва по-бавно и е по-продължително. Към тях се отнасят гладките мускули на стомаха, пикочния мехур и др. В тях подреждането на актиновите и миозиновите нишки не е равномерно, поради което не се наблюдава напречно набраздяване.
|
Независимо от разликите в строежа на различните типове мускули, при всички меха-низмът на мускулното съкращение е еднакъв и е свързан с взаимодействието на белтъците актин и миозин (Фиг. 15.2). Миозиновите нишки са по-дебели (имат диаметър около 10 nm), състоят се от молекули, който имат форма на дълга пръчка с две глобуларни образувания в единия край, наречени главички. В глобуларните образувания е концентрирана актин-свързващата способност на белтъка. Молекулите на миозина са подредени по такъв начин, че главичките са перпендикулярни на нишката и са разположени по спирала под ъгъл 1200 с период на повторение 42.9nm. Тънките изотропни нишки имат дебелина от 5 до 7 nm и дължина до 2.6 μm. Те се състоят от две преплетени вериги, изградени от подобни на перли актинови мономери. |
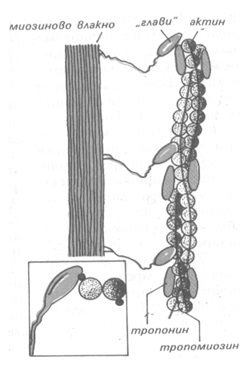
С актиновите молекули са свързани по-леките регулаторни белтъци тропонин и тропомиозин. На равни разстояния (около 40nm) по актиновите молекули са разположени кълбообразни тропонинови молекули. В образуващите се между актиновите вериги над-лъжни улеи се намират нишки от тропомиозин. Тази тропонин-тропомиозинова система представлява чувствителен към Са2+ йони преобразувател, управляващ съкратителните процеси. В състояние на покой те са така подредени, че да блокират участъка от актиновата молекула, който може да образува комплекс с главичките на миозиновите моле-кули. Когато в резултат на електричен импулс свободните Са2+ йони достигнат критична концентрация, локализирания на актиновата нишка тропонин-тропомиозинов комплекс претърпява конформационно изменение, което освобождава миозин-свързващия участък от молекулата на актина. Образува се комплекс между миозиновите мостчета и актиновата молекула и се развива мускулна сила, като се разгражда АТФ. При това се променя наклона на главичката на миозиновата молекула, миозиновата нишка се хлъзга между актиновите нишки и мускула се съкращава. При съкращение дължината на тънките и дебелите нишки не се променя, а те се плъзгат една между друга (Фиг. 15.3)
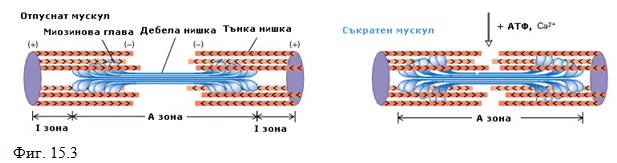
Процесът на съкращение се прекратява, когато възбудата на мембраната затихва, при което Са2+ йони се поемат отново от саркоплазмената мрежа.
През време на фазата на съкращение на мускула той съхранява чувствителността към нови дразнения. Продължително съкращение, предизвикано от следващи едно след друго импулси се нарича тетанично.
Мускулното съкращение като състояние на механична активност е два вида – изометрично и изотонично. Ако мускула е закрепен неподвижно, при възбудата му се повишава напрежението в него, но той не се съкращава. При това се отделя топлина. Такова съкращение се нарича изометрично. Ако мускула е закрепен подвижно, то при възбуда той се съкращава и енертията, необходима за съкращението се преобразува както в топлина, така и в механична работа Такова съкращение се нарича изотонично. Големината на силата, развивана от мускула при съкращение зависи от степента на припокриване на нишките, т.е. от силата е пропорционална на броя на потенциално възможните контакти на миозиновите мостчета с актина.
Топлината, която се отделя в мускула при единично изотонично съкращение можем да разделим на две части – топлина на активация Qa и топлина на съкращение Qc. Топлинният ефект на химичните процеси, които протичат при прехода на мускула от невъзбудено във възбудено състояние, представлява топлината на активация. Отделя се непосредствено след прилагане на дразнене върху мускула, преди появата на механична реакция. Големината и не зависи от механичната работа на мускула. Топлината на съкращение зависи от степента на съкращение. Ако големината на съкращението означим с X, Qc=a.X, където a е константа. Следователно 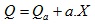
При всяко свиване на мускула той извършва работа A=P.X, където Р е механичното напрежение в мускула. Следователно изменението на вътрешната енергия на мускула ще бъде 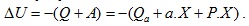 Тогава коефициентът на полезно действие η на мускула (отношението на полезната работа към промяната на вътрешната енергия) ще бъде
Тогава коефициентът на полезно действие η на мускула (отношението на полезната работа към промяната на вътрешната енергия) ще бъде 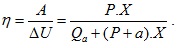 За различните организми този коефициент е различен, като за човека е приблизително 40%, за жаба 45%, а за костенурка 75%.
За различните организми този коефициент е различен, като за човека е приблизително 40%, за жаба 45%, а за костенурка 75%.

