

IV. РЕПЛИКАЦИЯ НА ДНК
Удвояването на молекулата на ДНК или на геномната РНК се нарича репликация и е основата при предаването на наследствената информация от поколение на поколение.
Репликация при прокариотите - полуконсервативен начин на репликация. Всяка новосинтезирана молекула ДНК съдържа една стара родителска и една нова дъщерна верига, синтезирана на базата на комплементарността на нуклеотидите.

Репликацията е ензимен процес - ДНК полимераза е ензимът на репликацията.
Репликон- всички пръстеновидни молекули ДНК, които са автономни единици на репликацията( молекула ДНК, която е способна на автономна репликация).
Типове репликация:
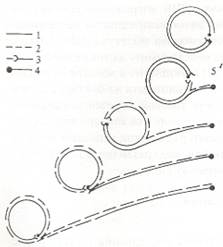
- Двупосочна репликация - Авторадиографските изследвания на Дж. Кернс (1963) на ДНК на E. coli показват, че репликацията при бактериите започва винаги от строго определен наследствено детерминиран участък на хромозомата, който се нарича начало на репликацията или точка на инициацията (О). Репликацията протича двупосочно в противоположни направления от точка О. Местата на хромозомата, до които е достигнала репликацията, се наричат точки на растеж или репликационни разклонения. При репликацията на ДНК на E. coli се образуват две постепенно придвижващи се по ДНК-матриците репликационни разклонения. След завършването на репликацията се получават две двуверижни пръстеновидни молекули, които отначало са свързани помежду си. При разделянето им единият пръстен временно се разкъсва. Този тип на репликация се наблюдава и при някои вируси, като например вирус SV-40, и при много плазмиди.
- Тип търкалящ се пръстен - Известна е още като размотаващо се руло. При този тип на репликация едната верига на двуспиралната пръстеновидна молекула на ДНК се разкъсва, а другата верига запазва пръстеновидната си форма. Върху освободените участъци на двете изходни вериги се осъществява полуконсервативна репликация, като се образува само едно репликационно разклонение. Интерес представляват особеностите на този тип репликация при малките фаги, като фаг fx-174, чийто геном се състои от едноверижна ДНК. Тя служи за матрица, върху която се синтезира комплементарна на нея верига. В резултат на това се получава двуверижен междинен продукт, наречен репликтивна форма. Изходната матрица се бележи като "+" верига, а новата комплементарна на нея - като "-" верига. В следващите поколения за матрица служат както "-" веригите, така и "+" веригите. По-ефективни матрици са "-" веригите, поради което по механизма на търкалящ се пръстен се синтезират предимно "+" вериги, които се включват в главичките на вирусните частици.
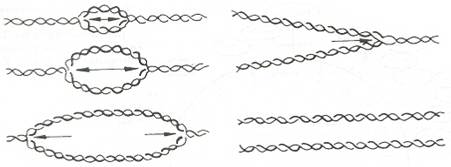
- Репликация на линейни молекули ДНК - Репликацията не започва от краищата на линейната молекула ДНК, а някъде в средата й (фиг. 25). Тя протича двупосочно, като началото на репликацията се открива лесно с електронен микроскоп, защото след започването на процеса на репликацията се образува подобен на око мехур, който се разширява към двата края на молекулата. Поради това, че мехурът достига единия край на молекулата по-бързо, отколкото другия, появява се Y-подобна конфигурация, която бързо преминава във V-подобна структура. Съдържащите се в 5' краищата на веригите на ДНК обърнати подреждания на нуклеотидите, които образуват палиндроми, имат важно значение за инициацията на репликацията.
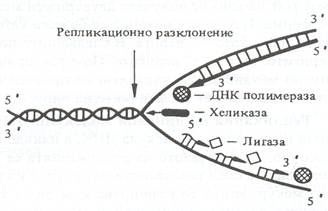
Генетична регулация на репликацията
Репликацията е под генетичен контрол и протича в три етапа - инициация, елонгация, терминация.
- Инициация - начало на репликацията на генома. Започва от синтезата на къси вериги РНК, които са праймър и синтезата им се катализира от ензима примаза. При E. coli процесът се регулира от гените dnaA, dnaC-polD, dnaH, dnaI и dnaP.
- Елонгация - растеж на ДНК веригата. Тя се катализира от ензими, синтезата на които се контролира от определени гени. Разкъсването на водородните връзки във всяко репликационно разклонение на ДНК се осъществява от ензима хеликаза. Получава се локално разделяне на двойната спирала на изходната молекула на ДНК, като репликационното мехурче постепенно се разширява двупосочно. Синтезираната върху ген ssb-1 свързваща белтъчина покрива едноверижните участъци ДНК, като ги предпазва от действието на намиращите се в клетката нуклеази. По-нататъшната елонгация на новосинтезираната верига при E. coli се осъществява от ензима ДНК-полимераза III, А-субединицата на която се кодира от ген dnaE. Функцията й е свързана и с активността на гените dnaB и dnaC. ДНК-полимераза III измества ензима примаза и използва съдържащите се в клетката дезоксирибонуклеозидтрифосфати за последователното удължаване на комплементарна на матрицата нова ДНК верига. Към 3'-края на всеки фрагмент празното пространство между два съседни фрагмента на Оказаки ензимът ДНК-полимераза I на E. coli, кодиран от ген pol A, присъединява няколко нуклеотида до достигането на 5'-края на другия фрагмент, от който отстранява РНК-закваската. Освен това ДНК-полимераза I изрязва неправилно включените в новата верига некомплементарни на матрицата нуклеотиди и ги заменя със съответстващи на старата верига нуклеотиди. Ензимът ДНК-лигаза, кодиран при E. coli от ген lig, свързва последователнокраищата на фрагментите, в резултат на което те стават по-дълги.
- Терминация - прекратяване на репликацията.
Репликация на ДНК при еукариотите
Разгледаните закономерности на репликацията при бактерии и вируси се отнасят и за еукариотните организми, но при тях възникват и специфични особености, свързани с равнището на тяхната организация. Молекулата на ДНК в хромозомата на еукариотите има линейно разположение. Репликацията на ДНК при тях се осъществява през S-фазата на интерфазата и протича полуконсервативно. За разлика от прокариотите при висшите еукариотни организми репликацията на ДНК започва от голям брой участъци по дължината на нейната молекула. Хромозомите на еукариотите съдържат множество независими участъци на репликация - репликони. Съседните репликони се разполагат по дължината на молекулата на ДНК един зад друг по двойки в тандем. Наблюдава се асинхронност при репликацията на хромозомите в различни тъкани на един и същ организъм и в рамките на една хромозома. Инициацията на репликацията в различни участъци на хромозомата започва по различно време. В един и същ момент отделни участъци са в различна фаза на репликация. Това показва, че съществува генетичен контролен механизъм, който определя в кои участъци на хромозомата да започне репликация и в кои да не започне. Следователно, инициацията на репликацията на хромозомите в клетките на животните става в определен ред.
От еукариотните клетки са изолирани три основни типа ДНК-полимерази: β - осъществяваща репликацията на ядрената ДНК; γ - репарационна полимераза на предизвиканите от ултравиолетови лъчи повреди на ядрената ДНК, и - осъществяваща репликацията на ДНК в митохондриите. ДНК-полимеразите и при еукариотите синтезират нови вериги ДНК само в посока 5'-3' края, като репликацията е прекъсната, защото се образуват фрагменти, подобни на фрагментите на Оказаки при бактериите.
Пресметнато е, че всяка клетка на бозайниците съдържа около 20 000 репликона. Те са по-къси при бързо растящите клетки с къс S-период в сравнение с репликоните при бавно растящите клетки с продължителен S-период. Скоростта на репликацията на ДНК при еукариотите зависи от плътността на свързване на хроматина в хромозомата. По-бърза репликация се наблюдава в пуфите, отколкото в дисковите и междудисковите пространства, тъй като пуфите са деспирализирани транскрипционно активни райони на гигантските хромозоми. Репликацията на хетерохроматиновите участъци протича асинхронно, защото те не се деспирализират едновременно целите, а на части. При определени условия отделни участъци от хромозомата реплицират многократно, докато през същия период репликацията на някои нейни участъци не завършва докрай. Върху броя на реплициращите се участъци в хромозомата влияе броят на наличните активни молекули ДНК-полимераза в клетката.
РЕПАРАЦИЯ НА ДНК
При повреда на някой участък на едната от веригите на ДНК в резултат на взаимодействието на разнообразани фактори, като радиоактивни лъчения, ултравиолетови лъчи или химични агенти, другата верига може да послужи като матрица за отстраняване на повредата и за възстановяване на изходното положение на нативната структура на ДНК, т.е. за репарация на структурата на ДНК в клетката. Познати са три основни механизма:
1. Фотореактивация - възстановяване на биологичната активност на повредените от UV лъчи на участъци от ДНК чрез въздействието на видимата светлина. Леталният ефект при облъчването на бактерии с ултравиолетова светлина се предизвиква поради образуването на димерен пръстен между два съседни остатъка на тимина, разположен в една от нуклеотидните вериги на ДНК. Това предизвиква локално нарушение на вторичната структура на двойната спирала на ДНК и потиска функцията на гена, в който е образуван димерният пръстен. Фотореактивацията се явява възстановяване на биологичната активност на повредените от ултравиолетовите лъчи молекули на ДНК чрез въздействие на видимата светлина с дължина на вълната от 300 до 500 nm. Фотореактивацията е ензимен процес, катализиран от изолирания от микроорганизмите ензим на фотореактивацията дезоксирибопиримидинфотолиаза, кодирана от ген phr. Субстрат на този ензим са димерите на пиримидиновите бази, с които на тъмно той образува комплекс. Той обаче не се свързва с неповредена молекула на ДНК. На светло комплексът се разпада, като от димерите се получават мономери. Видимата светлина осигурява необходимата енергия за ензимното разпадане на димерите. При мутантните бактерии E. coli с изменена структура на ген phr процесът на фотореактивация е блокиран, поради което при облъчването им с видима светлина тиминовите димерни пръстени на ДНК, предизвикани от облъчването с ултравиолетови лъчи не изчезват.
2. Ексцизионна репарация- отстраняване на повредени участъци на ДНК чрез изрязване. Ексцизионната репарация включва следните етапи: а) откриване на димерите; б) инцизия, т.е. надрязване на веригата на ДНК близо до димера; в) ексцизия, т.е отстраняване на димера; г) ресинтезиране на ДНК и д) възстановяване на непрекъснатостта на репарираната верига. Откриването на димерите в двуверижната ДНК се осъществява от ензима ултравиолетова ендонуклеаза. Тя разпознава не само димерите на тимина, но и всички най-разнообразни локални нарушения в двуспиралната структура на ДНК. Инцизията, т.е. надрязването на повредената верига на ДНК от 5'-края до димера, се извършва също от тази ендонуклеаза, включваща две субединици, синтезата на които при E. coli се детерминира от гените uvrA и uvrB. Мутациите на тези гени предизвикват повишена чувствителност на бактериите към ултравиолетовите лъчи. Ексцизията, т.е. изрязването на пиримидиновите димери от молекулата на ДНК, се извършва от ултравиолетовата екзонуклеаза. Ресинтезирането на ДНК се осъществява върху незасегнатата единична верига. При мутантите polA, recA, recB и recC репарационният синтез не завършва и при ниски дози на облъчване, защото настъпилите мутации са нарушили синтезата на необходимите ензими. Репарационният синтез на ДНК се осъществява от кодирания от ген polA ензим ДНК-полимераза I. Възстановяването на непрекъснатостта на репарираната спирала на ДНК протича под въздействието на ензима полинуклеотидлигаза, който при E. coli се кодира от ген lig. Чувствителните към повишена температура мутанти по ген lig не могат да завършат процеса на ексцизионната репарация при неподходяща температура.
3. Пострепликационна репарация- нарича се още рекомбинационна репарация и се наблюдава и при еукариотите. Наблюдава се, когато съдържащите тиминови димерни пръстени или други повреди участъци в молекулата на ДНК не се репарират преди S-периода и преди да започне процесът на репликация на ДНК. Тогава чрез полуконсервативен тип на репликация върху веригата ДНК с тиминови димерни пръстени се синтезират фрагменти нова верига ДНК, комплементарни на нормалните участъци на майчината верига, но с пролуки срещу димерните пръстени на матрицата, т.е. празни пространства, наречени гепове. Такива пролуки между новосинтезираните фрагменти ДНК се образуват при облъчване с ултравиолетова светлина. Тези пролуки изчезват при инкубацията на клетките в хранителна среда, което показва че е настъпила репарация. Но пострепликационна репарация не настъпва при rec-мутантните бактерии E. coli. Мутациите на rec-гените, кодиращи процеса на рекомбинацията, възпрепятстват репарацията. Въз основа на това се смята, че този тип репарация настъпва нормално чрез рекомбинация с комплементарни на майчината верига неповредени участъци ДНК вериги.
При нарушаване на механизмите на репарация на ДНК при човека и бозайниците могат да се появят тежки наследствени заболявания.
IV.ТРАНСКРИПЦИЯ
Процесът на презаписване на генетичната информация от молекулата на ДНК в РНК молекули се нарича транскрипция. Това е първи етап от функционирането на гена. Първичната структура на РНК е точно копие на първичната структура на кодиращия ген. Молекулата на РНК е полинуклеотидна верига близка по състав и строеж с ДНК. Тя бива 3 вида:
- информационна РНК (иРНК);
- транспортна РНК (тРНК);
- рибозомна РНК (рРНК).
Най-голямо количество в клетката е рРНК над 80%, а тРНК е 15%. ДНК служи като матрица за синтезирането на трите типа молекули РНК и те са копие само на дясната верига ДНК. Ензимът, който осъществява синтезата на РНК върху ДНК матрицата се нарича РНК полимераза. При еукариотите транскрипцията е много сложен процес с множество РНК полимерази.
Етапи на транскрипцията
Транскриптон - участък от молекулата на ДНК, който се презаписва като една молекула РНК.
1. Инициация - разпознаването на специфични подреждания на нуклеотидите в молекулата на ДНК. Наричат се промотори. Започва с локална денатурация на двойната спирала на ДНК. След това се създават комплекси между РНК полимеразата и ДНК. При наличието на нуклеозидтрифосфати в клетката към комплекса ДНК-РНК-полимераза се присъединява нуклеотид и започва синтезата на РНК. Това е началото на синтезата на РНК.
2. Елонгация - удължаване на полинуклеотидната верига на РНК. Успоредно с преместването на РНК полимеразата по дължината на РНК се присъединяват нови нуклеотиди. Синтезираната РНК верига временно е свързана с ДНК веригата под формата на хибриден участък след което се разделя и двойната верига на ДНК се възстановява.
3. Терминация - завършване на процеса на транскрипцията , при който се отделя РНК полимеразата и процесът приключва. Това се дължи на специфични подреждания на нуклеотидите в молекулата на ДНК при достигането на които РНК-полимеразата преустановява своята функция. Най-често нуклеотидните последователности на терминатора съдържат палиндрома, след която следват определен брой аденинови остатъци, след която РНК-полимеразата рязко забавя придвижването си и в разположения в съседство участък, съдържащ поли-А последователност, се отделя от веригата на ДНК. Съществуват и терминиращи сигнали, които се разпознават само в присъствието на белтъчните фактори (ро-фактор, капа-фактор, Nus-белтъчина и др.).За терминацията при еукариотите е известно много малко. Изглежда при тях тя не е много точен процес, защото синтезата на РНК не спира в определена точка, а в участък, който при ДНК е богат с тимин.
Зреене на РНК (Процесинг)
Новосинтезираните в ядрото молекули РНК се наричат хетероядрена РНК (хяРНК), част от която остава в ядрото, а останалата като пре-иРНК претърпява зреене и се превръща в иРНК. Зреенето протича чрез деградация, при която специфични ензими изрязват неактивните участъци на първичния транскрипт (интроните), а остават активните участъци (екзоните). Те се съединяват помежду си чрез ензима лигаза. Този процес се нарича сплайсинг (снаждане). Същевременно зреенето протича и с химична модификация (ацетилиране, металиране и фосфорилиране), при която в началото на иРНК се прикрепва една химична групировка, наречена кепе. В края на и РНК остава поли-А участък, наречен опашка, която е от значение за кратността на транслация.

