

VIII.Регулация активността на гените
- Всяка клетка съдържа пълния набор генетичен материал, но не всички гени се подлагат на транскрипция. Само 10-20% от гените са активни.
- В клетките има генетични механизми, които регулират активността на гените избирателно.
Основна роля в тази регулация играят ензимите. - Конститутивни ензими - постоянно се синтезират в клетката.
- Адаптивни ензими- синтезират се само когато са необходими в клетката. Те катализират катаболитните процеси. Субстратът на ензимите индуцира техния процес, който се нарича ензимна индукция.
- Анаболитните ензими се регулират чрез репресия - синтезирането на излишно количество от субстрата спира синтезата.
Регулация на генната активност при прокариотите - Осъществява се чрез негативна индукция и негативна репресия.
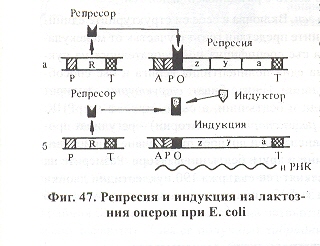
Негативна индукция - модел на лактозния оперона, който кодира синтеза на ензима лактаза при Ешерихия коли. Френските учени Жакоб и Моно, носители на Нобелова награда, при опити с E. coli проучиха лактозния оперон и създадоха модел за генетична регулация на синтезата на белтъчините или т.нар. модел на оперона. Лактозният оперон кодира синтезата на ензима лактаза за изграждане на субстрата лактоза, който E. coli използва когато в средата липсва предпочитаната от нея глюкоза. В изграждането участва система от ензими, които се кодират от група последователно подредени структурни гени (z, y, a). Те се транскрибират заедно като обща иРНК, а генът който отключва тази транскрипция е ген-оператор (о). Пред него се намират промотор (р) и участък (А) за захващане на белтък активатор. Цялата тази система от структурни и регулаторни гени се нарича оперон. Той изпълнява една функция. Операторът се включва и изключва от ген-регулатор със собствен промотор (р) и терминатор (т), който кодира синтезата на белтък - активен репресор. Репресорът може да блокира оперона като разпознава и се свързва с оператора. Същевременно лактозата може да се свързва с репресора и по този начин отключва действието на оперона. Lac-оперонът действа по следния начин. Когато в клетката липсва лактоза, репресорът е свързан с оператора и оперонът е блокиран. Не се кодира синтезата на ензим за разграждане на лактазата. При постъпване на субстрата лактоза в клетката, тя разпознава и се свързва с репресора, операторът се отключва и се кодира синтезата на ензимите, които разграждат лактозата до нейното пълно изчерпване от клетката. Ако мутират ген-операторът и ген-регулаторът, ензимите се синтезират конститутивно. Мутациите в областта на оперона доказват, че трите структурни гена са с общ оператор и терминатор, като транскрипцията в посока от ген z към ген а. Тъй като при Lac-оперона се синтезира активен репресор, който блокира транскрипцията, а постъпилият в клетката субстрат инактивира репресора и индуцира транскрипцията, този процес се нарича негативна индукция.
Освен по този индукционен път Lac-оперонът се активира и по втори път - чрез САР-белтък (белтъчина, активатор на катаболитните гени). Тя се свързва с А-област (при определени обстоятелства) и чрез промотора (Р) и ензима РНК-полимераза се активира транскрипцията.
Този първи модел на функциониране на гените трудно се доказва при еукариотите, но са известни макар и само единични случаи за регулиране на генното действие при тях по оперонния модел.

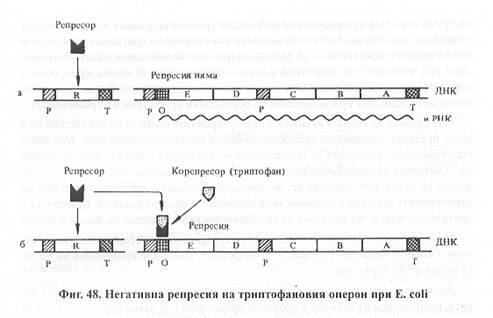
Негативна репресия - когато в клетката се натрупа излишък от субстрата, той се свързва с репресора, активира го и го превръща в активен репресор той се свързва с ген-оператора и блокира транскрипцията. Пример за такъв тип регулация е анаболитният триптофан-оперон, кодиращ синтезата на аминокиселината Триптофан. Той има същото устройство с някои различия: съдържа 5 структурни гена, 2 промотора, а регулаторният ген (R), кодира синтезата на неактивен репресор (апорепресор). В такъв случай операторът е свободен, оперонът отключен и се извършва транскрипция върху структурните гени. Така в клетката се синтезира триптофан. Когато в клетката се натрупа излишък, той се свързва с репресора, активира го и го превръща в активен репресор (холорепресор). Той се свързва с ген-оператора и блокира транскрипцията. При изчерпване на триптофана в клетката, холорепресорът се разрушава. Апорепресорът има друга конфигурация и не може да се свързва с оператора. Така в клетката се регулират автономно метаболитните процеси според нуждите й, без да е задължително регулацията на оперона да е строго индуктивна или репресивна. Един оперон може да отговаря и на едните и на другите сигнали.

Регулация на генната активност при еукариотите
Регулаторните механизми при тях избирателно активират определени групи гени. Характерна е групова регулация на цели блокове гени. Осъществява се на три нива:
- Претранскрипционно - осъществява се от хистоните, които неспецифично репресират гените и нехистоновите белтъчини, които специфично ги активират.
- Транскрипционна регулация - протича в оперони
- Посттранскрипционна регулация - роля играят интроните. Един ген може да кодира няколко вида полипептидни вериги в зависимост от това как е протекъл процесингът.
Доказано е, че един ген може да кодира повече от един вид полипептидна верига, което зависи от това как е протекъл процесингът. В някои случаи може да се изрежат едни интрони от пре-иРНК и да се получи един вид иРНК, в други случаи да се изрежат други интрони и да се получи друг вид иРНК, кодираща съвсем друга полипептидна верига. Така пре-иРНК на хормона калцитонин (от щитовидната жлеза) и на специфичен пептид на хипофизата е еднаква, но процесингът протича по различен начин, поради което и крайните продукти са различни. Същевременно иРНК при отделните видове еукариоти има различна продължителност на живот - от няколко минути до няколко дни (в сперматозоидите), а понякога месеци и години (в спорите).
Възможна е е генна регулация и при транслацията. Така в неоплодените яйцеклетки количеството иРНК е голямо, но транслацията е репресирана и тя започва едва след оплождането. В цитоплазмата на яйцеклетката има белтъчни фактори, които отключват не само транслацията, но и транскрипцията.
Основният механизъм на генната регулация при еукариотите е транскрипционният. Две групи механизми регулират експресията на гените:
Модулиращи - осъществяват количествен контрол според нуждите на клетката.
Алтернативни - осъществяват контрола на принципа пълно включване и пълно изключване.
Цитоплазмена нехромозомна наследственост
Наследствените структура в цитоплазмата се нарича плазмон, а самите наследствени елементи плазмогени.
Пластиди- плазмогени, които определят пъстролистността. Състоят се от ДНК, РНК и специфични белтъчини. Те се самовъзпроизвеждат.
Митохондрии- състоят се от ДНК, РНК, белтъчини, липиди, фосфатиди и ензими. Самовъзпроизвеждат се.
Взаимодействие между геном и плазмон
Геномът оказва влияние върху фенотипното проявление на плазмона.>Симбионти- чужди единици в цитоплазмата, които могат да предават нехромозомна генетична информация.
Епизоми- молекули ДНК в цитоплазмата, които могат да се включат в хромозомите и да проявяват свойствата на хромозомна наследственост.
Наследственост и среда
Фенотипното проявление на признаците е резултат от взаимодействието на генотипа и на средата.
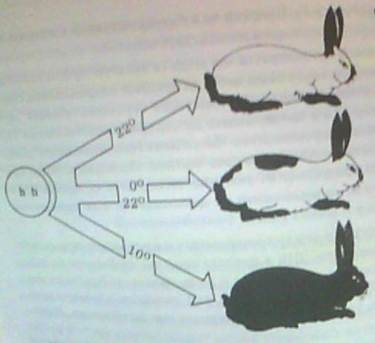
За установяване относителната роля на генотипа и средата се използват два метода:
- Организми с различни генотипове се отглеждат в еднакви условия на средата. Така се проследява влиянието на наследствеността
- Организми с еднакъв генотипи се поставят при различни условия на средата за да се отчете влиянието на средата върху фенотипа. Такива животни са само еднояйчните близнаци.

