

I.МЕХАНИЗМИ ЗА УНАСЛЕДЯВАНЕ НА ПРИЗНАЦИТЕ
В основата на развитието на генетиката стоят установените от Грегор Мендел закономерности, известни като Менделови закони.
Монохибридно кръстосване
- Първи закон на Мендел: при кръстосване на организми с противоположни признаци потомството в първо поколение е еднообразно. При съешаване на кръстоските от първо поколение (F1) помежду си във второто поколение (F2) се получава разпадане в съотношение 3:1, т.е. 75% от потомството е с доминантния и 25% - с рецесивния признак.
- Втори закон на Мендел: При кръстоските от първо поколение настъпва разпадане на признаците в точно определено отношение (3:1) за всеки признак.
- Закон за чистота на гаметите: При гаметогенезата през мейозата, поради случайното разпределяне на наследствените фактори, в гаметите по теорията на вероятностите може да попадне само фактор А или фактор а.
- Трети закон на Мендел: при кръстосването на организми, които се различават по два или повече признака, тези признаци се унаследяват независимо един от друг, като се комбинират във всички възможни съчетания.
- Анализиращо монохибридно кръстосване. Мендел използвал метода на анализиращото (обратното) кръстосване за разграничаване хомозиготните от хетерозиготните кръстоски в F2. При кръстосване на хетерозиготни организми от F2 (Аа) с рецесивни хомозиготни от изходната родителска форма (аа) се получава разпадане в съотношение 1 (Аа) : 1 (аа), а при кръстосване на хомозиготни с доминантния признак (АА) със същата родителска форма всички припло¬ди са хетерозиготни (Аа) и с доминантния признак.
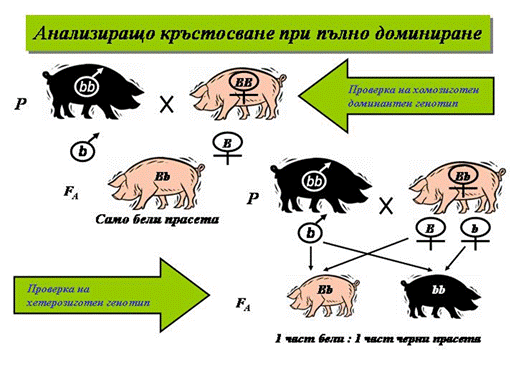
Фиг. 1. Анализиращо кръстосване
Дихибридно кръстосване
При дихибридното кръстосване в F2 се наблюдава разпадане на признаците в съотношение 9 черношарени без рога (с двата доминантни признака) : 3 черношарени с рога (с единия доминантен и единия рецесивен признак) : 3 червеношарени без рога (с другия доминантен и другия рецесивен признак) : 1 червеношарено с рога (с двата рецесивни признака). Съотношението 9 А-В- : 3 А-bb : 3 aaB- : 1 aabb може да се трансформира по следния начин: (3 А- : 1 аа) х (3 В- : 1 bb) = (3:1)2 Разпределението на генотиповете в F2 е 1 ААВВ : 2 ААВb : 1 AAbb : 2 AaBb : 4 AaBb : 2 Aabb : 1 aaBB : 2 aaBb : 1 aabb. Това е възможно само при независимото разпределение на гените в следващото поколение.
Интермедиерно унаследяване - При кръстосване на бели и черни андалузки кокошки кръстоските в F1 са сиви. В F2 25% са бели, 50% - сиви и 25% черни, т.е. признаците се разпадат в съотношение 1:2:1. Съотношението по фенотип напълно съвпада с това по генотип. В F2 потомството на белите кокошки е само бяло, на черните - черно, а при сивите се наблюдава разпадане, както в F1 в съотношение 1:2:1, което показва, че те са хетерозиготни по генотип.
Свръхдоминиране. Това е тип на взаимодействие между алелите на един ген, при който величината на признака в хетерозиготите превъзхожда този в хомозиготите - АА < Aa > aa. Потомството от F1 превъзхожда и двете изходни родителски форми по обща жизнеспособност, растеж и развитие, плодовитост, здрава конституция, продуктивност, устойчивост към болести. Смята се, че свръхдоминирането лежи в основата на хетерозиса при организмите.
Кодоминиране. При него алелите на даден ген проявяват еднаква степен на доминантност. По този тип се унаследяват кръвните групи при човека и животните - АВ и А1А2 съответно.
Взаимодействие на гените
- Комплементарно унаследяване. Комплементарният тип на взаимодействие е характерен при унаследяване на различната форма на гребените при кокошките, която се детерминира от 2 неалелни гена в две алелни състояния. Доминантният ген R детерминира розовиден гребен, а рецесивният r - листовиден (прост) гребен. Доминантният ген Р детерминира граховиден, а рецесивният р - листовиден. Гените R и P не са алелни и са разположени в различни хромозоми. При кръстосване на кокошки от породата Виандот с розовиден гребен (RRрр) с петли от породата Корниш с граховиден гребен (rrРР) в F1 се получават дихетерозиготни индивиди с ореховиден гребен (RrPp), който не се среща в родителските форми (първи закон на Мендел). В F2 разпадането е както при дихибридното кръстосване - 9 (R.P. - ореховиден) : 3 (R.pp - розовиден) : 3 (rrP. - граховиден) : 1 (rrpp - листовиден). Новата форма - ореховиден гребен е резултат от взаимодействието между доминантните неалелни гени R и P.
- Епистаза - взаимодействие между неалелни гени, при което единият ген подтиска фенотипната изява на другия ген. Потискащият ген е епистатичен, а потиснатият - хипостатичен. Доминантна епистаза се наблюдава при кръстосването на сиви и алести коне, на кокошки Бял легхорн с Бял плимутрок и др. Генотипът на изходните сиви коне се означава с ААВВ и на алестите - с ааbb. Кръстоските от първо поколение са сиви с генотип AaBb, а в F2 се наблюдава разпадане по фенотип в съотношение 12:3:1. От общия брой на приплодите 12/16 с генотип A-B- и A-bb са сиви, 3/16 (aaB-) са черни и 1/16 (aabb) - алести. Доминантният ген А в хомозиготно или хетерозиготно състояние действа епистатично срещу ген B и ген b. При всички комбинации, в които действа ген А (9 A-B- и 3 A-bb), се проявява сив цвят на косъма. Кончетата с генотип ааВ- са врани (черни), освободеният от супресията ген В проявява своето действие. Приплодите с двата рецесивни гена са алести.
- Полимерия - тип на взаимодействие между неалелни гени, при който развитието на признака зависи от броя на гените, които го детерминират. Гените от този тип се наричат полимерни, явлението - полимерия, а признаците - полигенни. Най-често това са количествените признаци - продуктивност, устойчивост и др., които се детерминират понякога от 100-200 гена. Полимерните гени се означават с един и същ символ, но с различен индекс (А1А1А2А2А3А3...). Класически пример за полимерия е унаследяване цвета на зърната на пшеницата по Нилсон Еле. При кръстосване на червени и бели зърна в F1 всички хибриди имат средно интензивен интермедиерен цвят. В F2 се наблюдава разпадане в съотношение 15:1, като 15 части са с различно интензивен цвят, в зависимост от броя на доминантните гени в генотипа, а 1/16 са бели, защото съдържат само рецесивни гени. Разпадането по генотип е 1:4:6:4:1.
- Трансгресия. При взаимодействието на полимерни гени може да се прояви явлението трансгресия. При нея отделни генотипове превишават или отстъпват по качествата си изходните родители. Затова тя бива положителна и отрицателна. Трансгресията се проявява, когато по-голям брой гени участват в регулирането на признака, но вероятността да се съберат в хомозиготно състояние всички доминантни гени в даден генотип е малка. Трансгресията се различава от хетерозиса по това, че при хетерозиса цялото потомство превъзхожда изходните родители, но в следващите поколения настъпва непрекъснато разпадане. На базата на трансгресията може да се провежда трансгресивна селекция за повишаване продуктивността и устойчивостта на животните към болести.
- Плейотропия. Понякога един ген влияе върху развитието на два или повече признака на организма. Явлението, при което един ген влияе върху няколко признака, се нарича плейотропия (от гръцката дума pleistos - множествен). Плейотропното действие на гените е свързано с това, че синтезираният под контрола на даден ген ензим влияе върху няколко биохимични реакции, които детерминират в една или друга степен (като основни или вторични реакции) развитието на признака. Колкото по-рано се проявява действието на плейотропния ген, толкова по-голям брой признаци могат да бъдат засегнати от него. Генът за жълтия цвят на космената покривка при лабораторните мишки, който в хомозиготно състояние е летален, предизвиква промени в обменните процеси, с което е свързано понижаването на плодовитостта и ранното им затлъстяване. При говедата от породата Декстер доминантният ген, който предизвиква хиперфункция на хипофизата, влияе върху пропорциите на телосложението и някои физиологични признаци, като понижен обмен, повишена скорозрелост и способност за угояване и има летално действие.
- Гени модификатори. По-голяма част от признаците се детерминират от един главен ген или олигоген и от различен брой второстепенни гени, които усилват или отслабват действието на олигогена. Такива гени се наричат гени модификатори. Например гени модификатори определят размера на белите петна по тялото на Черношареното и Айрширското говедо, а също и на непигментираните пръстени около очите на Херефордското говедо, които са причина за рак на очите при силно слънчево облъчване.
- Генен баланс и генотипна среда. Процесът на развитието на признаците е резултат на сложно взаимодействие между алелни или неалелни гени, усилване или отслабване действието на главния ген под влияние на гените модификатори или на плейотропното действие на някои гени. Има данни, че развитието на признака може да зависи от целия генотип на организма. Въз основа на това са създадени понятията генен баланс, т.е. съотношението при взаимодействието на всички гени, които влияят върху развитието на признака, и генотипната среда, т.е. комплексът от всички гени в организма, в който се проявява действието на даден ген. Генният баланс може да бъде променен от различни фактори: мутации, кръстосване на индивидите, подбор, отбор и др., а тази промяна оказва влияние върху развъдната работа в животновъдството.

