

II. Хромозомна теория за наследствеността
Хромозомите са самопроизвеждащи се постоянни структури на клетъчното ядро при еукариотите. Те се отличават със сложна морфологична, функционална организация и индивидуалност. Хромозомите са материалните носители на наследствената информация.
Хромозомите представляват плътни компактни структури, които се образуват при крайна степен на спирализация на хроматина. Могат да се наблюдават с оптичен микроскоп по време на метафазата на клетъчното делене. Всеки вид организми има свои отличителни белези,следователно - своя специфична генетична програма, която е поместена в специфичен брой хромозоми със свое характерно устройство.
Броят, формата и размерите на компактните хромозоми в телесните (соматични) клетки от един вид са негова характерна отличителна особеност. Тя се нарича кариотип. Броят на хромозомите, наречен хромозомен набор, при нормални телесни клетки се състои от двойки еднакви хромозоми (хомоложни хромозоми), тъй като в зиготата са се събрали бащиният и майчиният хромозомен набор, съответно от сперматозоида и яйцеклетката. Двойният хромозомен набор се нарича диплоиден (2n). В половите клетки хромозомният набор е единичен хаплоиден, (n).
По време на интерфазата, предхождаща същинското клетъчно делене, се извършва процесът репликация. Удвоява се количеството на генетичния материал. Той се намира под формата на хроматин, който се състои от дълги и тънки нишки(хроматинови нишки), изградени от ДНК и ядрени белтъци. Тези нишки са силно преплетени едни в други. Тогава започва процес на спирализацията им и така те се разплитат едни от други. Чрез напредване на процеса се стига до етап на свръхспирализация и образуване на хромозомите.
Всяка хромозома се състои от две части, които се наричат хроматиди. Те са напълно идентични помежду си и са резултат от удвояването на ДНК чрез процеса репликация. Всяка хроматида следователно съдържа една молекула ДНК. При клетъчното делене те се отделят една от друга. Всяка хроматида се обособява като хромозома за съответната дъщерна клетка. Мястото на свързване на двете хроматиди се означава като центромер. Там се намира и т.нар. първично прищипване , което разделя хроматидите на две рамена. Рамената могат да са с еднаква или различна дължина. Крайните участъци на рамената се наричат теломери. При някои хромозоми се наблюдава и т.нар. вторично прищипване, което се означава като организатор на ядърцето. Чрез вторичното прищипване се формира участък от хромозомата - сателит. В него са съсредоточени гените за рРНК, които образуват ядърцето на интерфазното ядро.
Размерът на рамената, броят и местоположението на вторичните прищипвания, както и формата и големината на сателита са специфична морфологична характеристика на всяка хромозома.
В зависимост от местоположението на центромера и дължината на двете рамена на хромозомата, разделяме хромозомите на :
- Метацентрични - двете рамена са почти еднакви по дължина , /p=q/.
- Субметацентрични - двете рамена се различават по дължина, като по-дългото рамо се означава с “q”, а по-късото рамо с “р”, /p<q /.
- Акроцентрични - центромерът е разположен почти в единият край на хромозомата, поради което едното рамо е много по-дълго от другото, /p<<q /.
- Телоцентрични - центромерът е разположен в края на хромозомата, в теломерния участък, поради което едното рамо липсва, т.е. има само едно рамо. Такива хромозоми не се наблюдават при човека, но са характерни за други животински видове.Телоцентричните хромозоми са обикновено нестабилни, поради което често се съединяват с други хромозоми, формирайки по този начин дицентрични формации, което довежда до усложнения при следващите деления.
При специфична активност на дадени гени се наблюдава раздуване на определени хромомери в съответния район на гигантските хромозоми. Такава раздута хромомера се нарича пуф или пръстен на Балбиани. Пуфите в политенните хромозоми на Chironomus се различават по форма и размери през отделните стадии на развитието. Смята се, че пуфите са външен израз на активността на гените на хромозомно равнище. Те са тъканно специфични, т.е. в различните тъкани са различни.
Четковидни хромозоми.
"Четковидните" хромозоми са стотици пъти удължени, силно деспирализирани хромозоми, чиято централна част се състои от плътни хромонеми (хромомери). На някои места от тях излизат извънредно дълги бримки, които придават на хромозомите вид на четки за миене на съдове. Смята се, че една молекула ДНК се е проточила от единия край на хромозомата до другия, като образува и хромонемите, и бримките. Докато ДНК в хромонемите е неактивна, в района на бримките тя е напълно деспирализирана, оголена и е място на активна транскрипция. Предполага се, че всяка бримка е отделен ген, съдържащ информация за синтезата_на една полипептидна верига.
Хромозомите са изградени от субстанцията хроматин. Хроматинът съдържа три вида молекули (ДНК, хистони, Кисели нехистонови киселини:
- Хистони- алкални нискомолекулни белтъчини, които стабилизират структурата на хромозомата и участват в регулацията на активността на гените. Известни са пет вида- Н1; Н2а; Н2b; Н3; Н4. Синтезират се в цитоплазмата и веднага преминават в ядрото където се свързват с новосинтезираните молекули ДНК . Това свързване не е видово специфично.
- Кисели нехистонови белтъчини имат регулаторна функция и са видово и тъканно специфични. Това са различните ензими, които регулиран процесите репликация, репарация , транскрипция и други.
Хроматинът бива два вида - еухроматин и хетерохроматин. Характерно за еухроматина е, че в него се съдържат почти всички протеин кодиращи гени. През целият живот на клетката този вид хроматин е в декондензирано състояние, като върху него постоянно се извършва транскрипция. Хетерохроматинът се разделя на два под вида:
- Конституитивен хетерохроматин
Конститутивният хетерохроматин се среща в кондензирано състояние в силно спирализираните участъци на хромозомите. Разполага се предимно в районите на центромера, организатора на ядърцето и теломерите в свободните краища на хромозомите. Той е генетично неактивен. Върху хетерохроматидните участъци почти не се синтезира РНК. В хетерохроматина се съдържа голямо количество сателитна ДНК, която е съвършено различна при видове от едно и също семейство. Хормоните не могат да активират хетерохроматина. Гените, които са разположени до хетерохроматиновите участъци, са подложени на частична инактивация (ефект на положението). - Факултативен хетерохроматин
Факултативният хетерохроматин се появява само през определени периоди от клетъчния цикъл, изключително в хромозомите на клетките от някои тъкани. Смята се, че той преминава от хетерохроматично в еухроматично състояние и обратно. Допуска се, че това се регулира от белтъчините в ядрото. - Факултативен хетерохроматин се образува при инактивацията на една от двете Х хромозоми през ранните стадии на формирането на зародишите от женски пол при бозайниците. Тъй като Y хромозомата е бедна на гени, разположените в Х хромозомите гени при женските бозайници са в двойна доза, а при мъжките - в единична. Това би трябвало да се отразява върху степента на развитието на свързаните с пола признаци при женските и при мъжките животни от даден вид. Установено е обаче, че свързаните с пола гени, разположени в хетероложната част на Х-хромозомата, функционират абсолютно еквивалентно при женските и при мъжките животни. М. Бар и Е. Бертрам (1949) открили недалеч от ядърцето на интерфазните ядра на соматични клетки на жени малко хроматиново телце. В клетките на мъже с нормален кариотип хроматиново телце, наречено полов хроматин или телце на Бар, липсва. При индивиди с хромозомни аномалии ХХХ, ХХХХ при жените и женските животни при бозайниците или XXY, XXXY при мъжете и мъжките животни броят на телцата на Бар в соматичните клетки е с единица по-малък от броя на Х-хромозомите. Според хипотезата на Лайън (1968) телцето на Бар се образува поради силна спирализация и кондензация на едната Х-хромозома през ранните етапи от ембриогенезата. Х-хромозомата е хетерохроматизирана и генетично неактивна, което се запазва при всички следващи клетъчни поколения. Разположените в нея гени се намират в състояние на репресия и през фазата на клетъчния цикъл върху тази Х-хромозома не се осъществява транскрипция на РНК. В клетката остава активна само едната Х-хромозома, независимо от кой родител произхожда - от майката или от бащата. Това се осъществява на базата на случайността. При следващи клетъчни деления обаче същата хромозома остава инактивирана за целия получен при деленията клетъчен клон. Хипотезата на Лайън за една активна Х-хромозома в клетките е проверена и правилността й е доказана при изследване върху генетичната регулация на биосинтезата на различни изоензими на глюкозо-6-фосфатдехидрогеназата.
Брой на хромозомите при някой видове животни:
- Носорог 84
- Гълъб 80
- Домашна патица 80
- Мускусна патица 80
- Пуйка 80
- Кокошка 78
- Куче 78
- Кон на Пржевалски 66
- Кон 64
- Муле 63
- Катър 63
- Магаре 62
- Коза 60
- Говедо 60
- Бизон 60
- Зебу 60
- Овца 54
- Бивол 50
- Шимпанзе 48
- Див заек 48
- Човек 46
- Домашен заек 44
- Мишка 40
- Лисица 38
- Котка 38
- Свиня 38
- Дива свиня 36
- Зебра 32
- Пчела 32-16
- Норка 30
- Азиатски елен 6
Скаченост на гените
Разположените в определена хромозома гени съставляват скачена група и се унаследяват съвместно. Установено е, че броят на скачените групи гени при различните видове винаги е равен на хаплоидния брой на хромозомите. Така при дрозофилата хаплоидният брой на хромозомите е 4, на който съответстват 4 групи скачени гени.
Като пример за скачено предаване на гените може да се използва случаят на кръстосването на дрозофили с черно тяло и нормално дълги крилца и дрозофили със сиво тяло и рудиментарни къси крилца. Сивият цвят на тялото на дрозофилата зависи от нормалния доминантен ген, който се означава с b+, а черният цвят - от мутантен рецесивен алел b (от black - черен). Дългите крилца на дрозофилата се регулират от нормалния доминантен ген vg+, а късите - от мутантен рецесивен алел vg (от vestigial - къси крилца). Понеже тези гени са локализирани в автозомите, няма значение кой от двата генотипа притежава майката и кой - бащата. При кръстосване на сиви дрозофили с къси крилца и черни с дълги всички кръстоски са сиви с дълги крилца, тъй като фенотипно се проявяват двата доминантни гена. Те са хетерозиготни и имат генотип b+vg/bvg+. При пълно скачване на гените в отделните хромозоми кръстоските F1дават два вида гамети: b+vg и bvg+, докато при независимото разпределяне на гените при дихибридното кръстосване се получават 4 типа гамети: b+vg+, b+vg, bvg+и bvg. При съешаването на мъжки хетерозиготни кръстоски от F1с генотип b+vg/bvg+и женски мухи, които притежават двата рецесивни признака с генотип bvg/bvg, т.е. при анализиращото кръстосване (фиг. 14, 1), в F250% от цялото потомство има сиво тяло и къси крилца с генотип b+vg/bvg и по фенотип е еднакво с единия от изходните родители. Останалите 50% са черни, с дълги крилца с генотип bvg+/bvg, като по фенотип са еднакви с другия родител. Биологичната роля на скачването на гените при унаследяването се състои в това, че като ограничава свободното комбиниране на наследствените фактори, то създава по-голяма устойчивост на признаците при дадения вид организми.
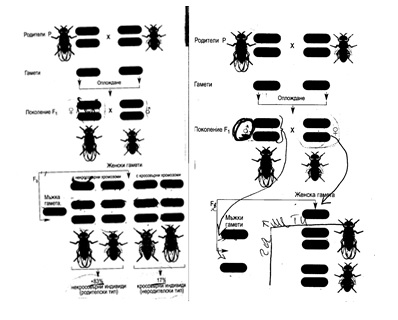
Фиг. 2. От ляво - Анализиращо кръстосване на мъжки хибрид от F1 с женски индивид, хомозиготен по рецесивния алел
От дясно - Анализиращо кръстосване на женски хибрид от F1 с мъжки индивид, хомозиготен по рецесивния алел
Генетична рекомбинация при еукариотите - Кросинговер
През профазата на първото делене на мейозата при т.нар. синапсис, хомоложните хромозоми - едната, идваща от майката и другата - от бащата, които са еднакви по форма и големина, се подреждат по двойки една срещу друга. След това всяка хромозома се разделя по дължина на две хроматиди, свързани чрез центромера. Четирите хроматиди на двете хомоложни хромозоми образуват т.нар. тетради.
Настъпва спирализация, при която хромозомите се увиват една около друга. В I профаза на мейозата се наблюдава своеобразно свързване (като бримка) между две несестрински хроматиди, наречено хиазма. Предполага се, че при този процес хроматидите от майчината и от бащината хромозома се разкъсват приблизително на едно равнище, т.е. на хомоложни локуси на мястото на хиазмата. Краищата на фрагментите срастват помежду си. Когато се съединяват хроматидите на двете хомоложни хромозоми, между тях настъпва обмяна на участъци. Едната част от така получените хроматиди, а по-късно хромозоми произхожда от майката, а другата - от бащата, при което гените на отделните хромозоми се прегрупират. Това явление се нарича кросинговер. Хиазмата е морфологичен израз на кросинговера. В резултат на кросинговера локализираните във фрагментите на една хромозома гени се предават на потомството скачени с гените, разположени в срастналия се в него фрагмент от другата хомоложна хромозома. При кросинговера настъпва рекомбинация на скачените гени, поради което се увеличават възможностите за комбинативната изменчивост при организмите.
По наличието на кросинговер се съди дали даден признак се контролира от един или от няколко гена. Но това може да се направи само при хетерозиготни организми. Когато гените са в хомозиготно състояние, дори ако настъпи прекръстосване на хромозомите, то не може да се открие, защото не довежда до промени в генотипа и фенотипа на потомството.
Кросинговер се среща при всички организми, при които се наблюдават мейоза и полова рекомбинация. Той протича при генетичен контрол, но се влияе от температурата на средата, възрастта, пола и др. При някои организми прекръстосването на хромозомите засяга само отделни участъци, а при други се разпространява равномерно по цялата дължина на хромозомата. Около центромерите и към краищата на хромозомите той се среща рядко. За различните двойки хромозоми честотата на повлияването на кросинговера не е еднаква. При отделните видове кросинговерът е ограничен от пола. Така при мъжките дрозофили и при женските копринени буби кросинговер изобщо не се наблюдава. Същевременно класически пример за кросинговер са опитите на Т. Морган с женски хетерозиготни дрозофили.
При кръстосването на сиви дрозофили с рудиментарни крилца b+vg/b+vg и черни с нормални крилца bvg+/bvg+ кръстоските от двата пола в F1 са хетерозиготни b+vg/bvg+ и по фенотип са сиви с нормални крилца.
При анализиращото кръстосване между хетерозиготните женски дрозофили от F1 с генотип b+vg/bvg+ и мъжки дрозофили, хомозиготни по двата рецесивни гена, т.е. черни с рудиментирани крилца, с генотип bvg/bvg, в F2 се получават 4 различни фенотипа. Те обаче не се разпадат в съотношение 1:1:1:1, както би съответствало на правилото за независимото разпределение. Двата фенотипа притежават признаците на изходните родители. Това показва, че гените, детерминиращи техните признаци, се предават по наследство скачени, т.е. през мейозата в хромозомите на половите клетки не са настъпили промени. Те се срещат в еднакво съотношение, по 41,5% от цялото потомство, или общо 83 вместо 100%.
Но едновременно с това 8,5% от приплодите имат черно тяло с рудиментирани крилца, т.е. двата рецесивни признака, а 8,5% са сиви с нормални крилца, т.е. те са носители на двата доминантни гена. Това е възможно само при рекомбинация на гените, вследствие на кросинговер между хомоложните хромозоми при узряването на половите клетки при женски дрозофили. Получените в резултат на кросинговера гамети и организми се наричат кросинговерни, а незасегнатите гамети от него - некросинговерни. При хетерозиготните женски дрозофили от F1 кросинговерни гамети са bvg и b+vg+, а некросинговерни - b+vg и bvg+. Възможността да се получат кросинговерни гамети показва, че скачването между дадените гени в хромозомата не е пълно.
Видове кросинговер
Тъй като гените в отделните хромозоми са много, в стадия на образуването на тетрадите при мейозата може да настъпи кросинговер на две или повече места между една и съща двойка хроматиди на хомоложните хромозоми. Тогава се говори за сложен кросинговер. Той бива възвратен, независим и некомпенсационен. Генетичните последствия при двойния възвратен кросинговер трудно се откриват, защото се възстановява изходното положение на двата гена и между тях не настъпва рекомбинация. За да се открие двоен кросинговер между една и съща двойка хроматиди, трябва да се направи анализ за унаследяването на трети ген, разположен по средата между мястото на двете разкъсвания на хроматидите. Когато второто прекъсване засяга останалите два хроматида на дадена тетрада и настъпва обмяна на фрагменти от четирите хроматида, говори се за независим сложен кросинговер. Ако втората хиазма се получи между една участвала в първото прекръстосване и друга, неучаствала хроматида, т.е. осъществява се обмяна между 3 хроматида, тогава се говори за некомпенсационен кросинговер.
Честотата на рекомбинацията зависи от разстоянието между гените. Между отдалечените скачени гени двоен кросинговер настъпва често, докато между близко локализираните гени се наблюдава рядко, защото вероятността за разкъсване на хроматидните нишки точно на това място е малка. Следователно, силата на скачеността е обратно пропорционална на разстоянието между гените. Предполага се, че при твърде големи разстояния на гените в хромозомата поради малката сила на скаченост между тях често настъпва кросинговер. Вследствие на сложността му се получава впечатление, че тези гени се комбинират свободно.
Степента на силата, която свързва скачените гени в една хромозома се измерва с процента на кросинговера между гените. Той се определя от честотата на рекомбинациите между скачените гени. Изчислява се по съотношението между броя на кросинговерните приплоди с нови комбинации на признаците и общия брой на приплодите при проведените опити с анализиращо кръстосване. Колкото по-висок е процентът на кросинговера, толкова по-слаба е връзката между скачените гени.
Опитните данни показват, че честотата на двойния възвратен кросинговер е по-ниска, отколкото се очаква теоритично. Това се свързва главно с явлението интерференция. Тя се характеризира със следното: когато се получи кросинговер в един участък на хромозомата, той потиска появяването на нов кросинговер в близките локуси на същата хромозома. Интерференцията е много разпространена. Потискащото й влияние се простира обикновено по дължината на едното рамо на хромозомата до центромера, където при силно изразена интерференция се появява само един кросинговер.
Интерференцията се измерва по честотата на рекомбинациите в потомството на хетерозиготни по няколко гена родители. Размерът на интерференцията за даден локус на хромозомата при определен генотип и константни външни условия е постоянна величина.
Генетична мярка за интерференцията е т.нар. фактор на съвпадението или коинциденцията. Тя представлява отношението между действително наблюдаваната и теоритично очакваната (при липса на интерференция) честота на двойния кросинговер. Разпределянето на кросинговера по дължината на хроматидите не е случайно, а се регулира от коинциденцията. Тя е обратно пропорционална на интерференцията. Стойността й варира от 0 до 1. При пълна интерференция липсва двоен кросинговер, поради което коинциденцията е 0. Когато няма интерференция, коинциденцията е 1. Колкото по-малка е стойността на коинциденцията, толкова по-малко двойни рекомбинации се получават.
Освен това известен е и соматичен кросинговер, при който се обменят фрагменти между хомоложни хромозоми на телесни клетки. Тъй като този процес протича при митотичното делене на клетките, той се нарича още митотичен кросинговер. Вследствие на този кросинговер при хетерозиготни организми се появяват клетки, които са хомозиготни по отношение на определен ген. Фенотипно това се проявява с възникването на мозаични признаци. При дрозофилата в резултат на соматичния кросинговер на фона на детерминирания от доминантния ген сив цвят се появяват жълти петна по тялото, предизвикани от рецесивен ген за жълтия цвят в хомозиготно състояние.
Генетика на пола
Полът е понятие в биологията, според, което група индивиди от даден биологичен вид, имащи възможност за полово размножаване, чрез специализацията в производството на определен вид полови клетки. Половете обикновено са два - мъжки и женски, като междуполовите биологични характеристики на половите жлези, половата физиология и/или ДНК полова хромозома се наричат хермафродитизъм.
Детерминирането на пола е биологичен феномен, който обуславя развитието на определен набор от признаци определящи пола при разделно половите организми. Най-общо детерминирането бива генетично, когато качеството и/или количеството на генетичния материал задава пола и негенетично, когато полът се определя от външни фактори.
Системата XX/XY за детерминиране е най-познатата и добре изучена, тъй като е представена при хората. При тази система женските индивиди имат две еднакви полови хромозоми(XX), докато при мъжките те са две различни (XY). Някои видове (в това число човекът) имат гена SRY носен от Y хромозомата, който детерминира развитието на мъжки пол; при други (като винената мушица) присъствието на две X хромозоми детерминира развитието на женски пол. Половите хромозоми X и Y се различават както по форма така и по размер за разлика от автозомите и се наричат алозоми. XX/XY системата се открива в повечето бозайници както и при някои насекоми.
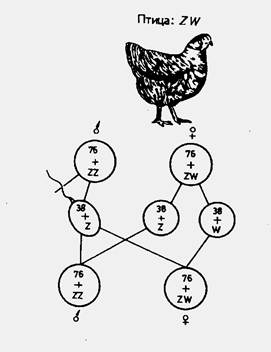
При птиците, пеперудите и влечугите с хетерогаметен женски пол е прието половите хромозоми да се отбелязват със Z и W. Мъжкият пол има ZZ полови хромозоми, а женският - ZW. Този тип на определяне на пола се нарича тип Abraxas (фиг. 3).
Определяне на пола при пчели
При пчелите, листните въшки и др. не се наблюдават полови хромозоми. Овогенезата протича нормално чрез мейоза, при което диплоидният брой хромозоми (2n = 32) се редуцира на n = 16. Сперматогенезата не протича чрез мейоза, а чрез митоза и те съдържат хаплоиден набор хромозоми (n = 16). При оплождане на яйцеклетката се получава диплоидна зигота (2n = 32), от която се развива пчела работничка или майка. При неоплождане на яйцеклетката, тя остава хаплоидна (n = 16) и от нея се развива търтей (фиг. 4), който е продукт на партеногенезис.
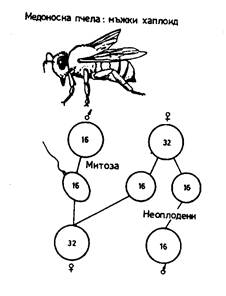
Фиг. 4. Определяне на пола при пчели.
Определяне на пола при насекоми
Скакалците и други видове организми с хетерогаметен мъжки пол нямат Y-хромозома (фиг. 5). Те имат два типа сперматозоиди. Едните от тях съдържат Х-хромозома и при сливането им с яйцеклетка, която винаги има Х-хромозома, се получава женски скакалец (ХХ). Другият тип сперматозоиди нямат полова хромозома и при сливането им с яйцеклетка се получава мъжки скакалец (ХО). Това показва, че и при тях Y-хромозомата не съдържа гени за определяне на пола.
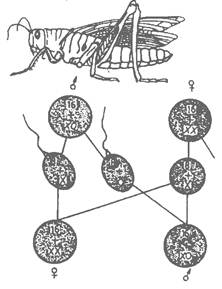
Фиг. 5. Определяне на пола при насекоми.
Гинандроморфизъм.
Гинандроморфизъм - полова аномалия при насекомите и някои птици, при която една част от тялото по генотип и фенотип е мъжка, а останалата - женска (фиг. 6). Той бива латерален, предно-заден, мозаичен и др. Механизмът на възникване е свързан с нарушения в ранния стадий на деление на зиготата, при което единият бластомер е с ХО-хромозоми и от него се развива мъжката половина, а другият с ХХХ - женската.
Има и други механизми за възникване на гинандроморфизъм. Въз основа на тези явления Бриджес създал балансовата теория за определяне на пола, според която всеки организъм е бисексуален и съдържа заложби за двата пола. Полът се определя по време на развитието на базата на баланса на мъжките и женските гени-детерминатори на пола.
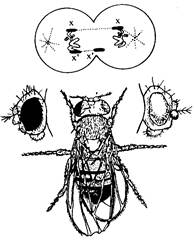
Фиг. 6. Латерален гинандроморфизъм при дрозофилата
Хермафродитизъм.
Хермафродитизъм - организми, които имат и двата типа полови органи, което води до промени във вторичните морфологични и физиологични признаци. Описаните 3 отклонения - гинандроморфизъм, интерсексуалност и хермафродитизъм, възникват при нарушаване на връзката между половата детерминираност, която се определя от генотипа и половата диференциация, под влияние на средата.
Фримартинизъм.
При бозайниците преопределянето на пола е трудно, но бисексуалната природа на зиготата създава условия за въздействие. При говедото през бременността се създават комуникации между кръвоносните съдове на близнаците. При разнополовите близнаци първи започват да функционират мъжките полови хормони и потискат развитието на яйчниците на женския зародиш. Затова след раждането бичетата се развиват нормално, а женските са безплодни и се наричат фримартини, а явлението - фримартинизъм
Хемизиготно състояние на гена.
Половите хромозоми се състоят от два участъка, единият от които е хомоложен за Х- и Y-хромозомите, а другият - хетероложен, т.е. различен. В хетероложния участък са локализирани както гените, определящи пола, така и други гени, които се предават на потомството, свързано с пола. Между хетероложните участъци никога не се наблюдава кросинговер, защото в тях няма алелни гени. Това състояние на гените се нарича хемизиготно. Между хомоложните участъци (фиг. 19, III до IХ) е възможен кросинговер, защото в тях се разполагат алелни гени, които са частично свързани с пола.
При реципрочната комбинация на кръстосване между хомозиготна женска дрозофила с бели очи и мъжка с червени очи в F1всички женски мухи имат червени очи, тъй като получават от бащата Х-хромозомата, която носи доминантния ген за червения цвят на очите. Същевременно всички мъжки кръстоски в F1има бели очи, защото те получават от майката Х-хромозомата с рецесивния ген за белия цвят на очите. Понеже Y-хромозомата не съдържа гени, които влияят върху цвета на очите на дрозофилата, фенотипно се проявява действието на рецесивния ген за белия цвят (хемизиготно състояние на гена). Този тип на унаследяване, при който майката предава признаците на синовете, а бащата - на дъщерите, се нарича унаследяване на кръст. В F2признаците се разпадат в съотношение 1:1:1:1, при което 25% от общия брой на индивидите са женски с бели очи, 25% - женски с червени очи, 25% - мъжки с бели очи и 25% - мъжки с червени очи. Понеже мъжките дрозофили имат само една Х-хромозома, опитите по унаследяване на признаци, свързани с пола, показват ясно в коя хромозома е локализиран генът, който определя дадения признак.
Регулиране на пола - същност и примери.
Полът, както всеки друг признак, е резултат от взаимодействието на генотипа и околната среда. Чрез промяна на условията на средата може да се въздейства върху нормалното развитие на пола. Преопределянето на пола е толкова по-лесно, колкото по-несъвършен е хромозомният механизъм за определяне на пола при видовете. Така например, сравнително лесно се преопределя полът при риби и амфибии. При отстраняване на половите жлези на жаба рудиментираните гонади се развиват и започва да функционира противоположния пол. При отстраняване на левия яйчник на кокошка се развива десният, а той започва да функционира като семенник и кокошката фенотипна придобива фенотип и поведение на петел.
При копринената пеперуда Астауров въздейства на неоплодени яйца чрез термичен шок - 46оС за 18 min и те се развиват партеногенетично чрез митоза вместо чрез мейоза. Потомството (100%) по генотип е като майката (ZW) - женски партеногенетичен клон. Явлението се нарича гиногенеза. В практиката, обаче са по-ценни мъжките буби, тъй като от тях се получава 25-30% повече коприна. Партеногенетично са получени и 100% мъжки пуйчета (ZZ). Получаването на мъжки партеногенетичен клон се нарича андрогенеза.
Направени са опити да се регулира пола при бозайниците чрез центрофугиране и разделяне на сперматозоиди с Х- и Y-хромозома на базата на различната им маса. Различията в електрическия им заряд откриват възможността за разделяне чрез електрофореза. Тези методи обаче не дават отчетлив резултат. По-перспективни за регулация на пола са имуногенетичните методи. Така при имунизиране на жена с антигени от сперматозоиди с Х- или Y-хромозоми се получават съответните антитела, които взаимодействат със съответните антигени от имплантираните зиготи и така се предопределя раждането на индивид с определен пол.
Възможно е в най-ранен стадий от развитието на ембриона да се постави ранна диагноза на пола на зародиша по наличието или отсъствието на телцето на Бар, което представлява инактивирана Х-хромозома. При мъжките то липсва.

