III. МОЛЕКУЛНА ГЕНЕТИКА
Молекулната генетика изучава на молекулно ниво генетичните механизми на регулиране на наследствеността и изменчивостта при живите организми. Обект на молекулната генетика са прокариотните и еукариотните организми. Най-подходящи за генетични изследвания са микроорганизмите, защото:
- имат кратък цикъл на размножаване и бърза смяна на поколенията
- просто устройство на генома
- евтино се отглеждат
- лесно достъпен е геномът за различни въздействия на околната среда
системата бактериофаг - бактерия, подходящ модел за изследвания на молекулно ниво.
Основните обекти са Ешерихия коли, червената плесен, бактериофагите и вирусът на тютюневата мозайка.
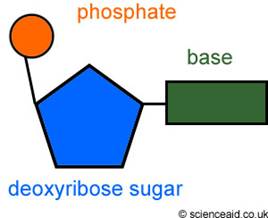
Нуклеотид - Молекула, съставена от азотна база (пуринова или пиримидинова), захар с пет въглеродни атома (рибоза или дезоксирибоза) и фосфатна група. Основна структурна единица на дезоксирибонуклеиновата (ДНК) и рибонуклеиновата киселина (РНК).
Молекулна структура и функции на ДНК
Структура на ДНК - Крик и Уотсън (1958 г.) за първи път построяват модел на двуспиралната структура на ДНК. Комплементарността на двете вериги ДНК осигуряват два пълни набора наследствена информаци, като всяка от тях може да бъде матрица при репликацията. Подреждането на базите в ДНК е първичната структура, която е и кодът на наследствената информация. То е произволно и същевременно специфично за всяка молекула ДНК.
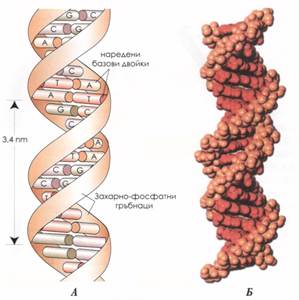
При двуспиралната ДНК самата спирална конфигурация представлява нейната вторична структура. Едноверижната молекула ДНК се среща при генома на бактериофагите. При тях липсва еквивалентност на базите.
Геномна РНК - това е едноверижна РНК, която се среща при някой вируси и фаги. В някои участъци може да има комплементарност, която изглежда като фиба и се образуват т.нар. палиндроми.
Денатурация, ренатурация и хибридизация на ДНК
- Денатурация на ДНК - разделят се двете комплементарни вериги на ДНК под влияние на висока температура.
- Ренатурация - свързване на денатурираните вериги чрез бавно охлаждане.
- Хибридизация - образуване на хибридни молекули ДНК чрез ренатурация на единични вериги ДНК, при което комплементарните им участъци се свързват. Останалите некомплементарни участъци остават несвързани и образуват хетеродуплекси.
Структура на ДНК при прокариотите
При тях се касае само за една хромозома, която най-често е пръстеновидна. При някои бактериофаги (Т2, Т4, Т7 и др.) генетичният материал е съсредоточен в двойноверижна ДНК с линейна форма. Добре е проучена наследствената структура на E. coli. Тя е двойноверижна пръстенообразна молекула ДНК, представена като ядрено телце - нуклеоид в бактерията. В разгърната форма има около 4 cm дължина. Тази дълга молекула трябва да бъде пакетирана така, че да се събере в малката бактерийна клетка и всеки ген да е достъпен за репликация и транскрипция. Предложен е модел за структурата на бактерийния нуклеоид (фиг. 20). ДНК е нагъната на 10-50 бримки и е суперспирализирана (1 спирала на 100 нуклеотида). Основата на всяка бримка е свързана от молекулата РНК, която като възел я поддържа, за да не се разгъва. Ако се третират възлите с ензима рибонуклеаза, бримките се разгъват, но суперспирализацията на молекулата остава. Тази структура позволява при транскрипцията да се транскрибират отделни бримки, без да се нарушава останалата структура на генетичния материал.
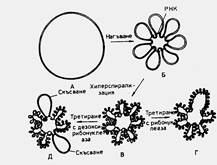
Фиг. 7. Структура на бактериалния нуклеотит (според Pettijohn & Hecht, 1974):
А - кръгово ненагъване на хромозома; Б - нагъната структура. Всяка бримка се осъществява при участието на молекули от РНК. Предполага се, че всяка хромозома образува 40-50 бримки, но на схемата са представени само 7 за улеснение на илюстрацията; В - свръхспирализация на ДНК-молекулата по дължината й; Г - при третиране с рибонуклеаза се разрушават мостовете, образуващи отделните бримки, но свръхспиралната структура се запазва; Д -при третиране с дезоксирибонуклеаза се получават скъсвания в една от веригите на ДНК, което води до изчезване на свръхспиралната структура само в съответната бримка
Генетична рекомбинация при прокариотите
Генетичната рекомбинация или кросинговър се наблюдава при всички ДНК-съдържащи организми без тези, съдържащи геномна РНК. Тя бива:
- обща (реципрочна) - обменят се участъци между две хомоложни хромозоми
- специфична (нереципрочна) - обменят се участъци от нехомоложни хромозоми.
При бактериите генетичната рекомбинация представлява трансформация, трансдукция и конюгация.
- Трансформация - част от ДНК на клетката донор се пренася и рекомбинира с ДНК на клетката реципиент. В следващите поколения този участък се предава заедно с ДНК на реципиента. Така бактериите придобиват нови свойства - вирулентност, устойчивост към лекарства и др.
- Трансдукция - пренасянето на фрагменти от хромозомата на една бактерия в друга бактерия чрез фаги и вируси. Тя бива - неспецифична, ако пренасяният фрагмент от донора е неспецифичен и може да се включи на всяко място в ДНК на донора и специфична - пренасяният фрагмент е специфичен и се включва на точно определено място. В някои случаи пренесената ДНК не се рекомбинира, а остава свободно в клетката и без да се възпроизвежда се предава само на едната дъщерна клетка - абортивна (незавършена) трансдукция. Пример за специфична трансдукция е пренасяне на локус gр1 от E. coli чрез бактериофаг. Този локус детерминира синтезата на ензима галактокиназа за разграждане на галактозата, която е основна храна на бактериите.
- Конюгация - парасексуален процес, при който генетичната информация се пренася чрез пряк контакт между две бактерии. Генетичната информация се пренася чрез полов фактор F, който F+ бактериите съдържат. Съществуват и F-бактерии, които са без полов фактор. Пренасянето се извършва в посока от F+ към F-.
Генетичната рекомбинация на бактерийните клетки се осъществява чрез участието на плазмиди. Плазмидите са автономно реплициращи се генетични структури в цитоплазмата вън от хромозомата на бактерийната клетка.
Плазмиди - Те представляват двуверижни пръстеновидни молекули ДНК. При много видове бактерии са описани различни типове плазмиди. Някои от тях могат да се включат чрез кросинговер в хромозомата на реципиентната клетка. Те се наричат епизоми. За наличието на плазмиди в бактерийните клетки обикновено се съди по детерминираната от тях функция. При еукариотните клетки също са наблюдавани аналогични образувания. Най-добре са проучени при Грам-отрицателните бактерии и преди всичко половият фактор F при Е. coli и R-факторите за пренасяне устойчивостта на бактериите към лекарствени средства. Добре проучени са и колициногенните фактори (col), съдържащи гени, които детерминират синтезата в реципиентната клетка на колицини, явяващи се токсини с летален ефект при колибактериите. Установено е, че половият фактор F и някои фактори R са твърде сходни помежду си и в много отношения те са аналогични с умерените фаги.
Плазмидите имат сравнително малки размери - от 1 до 200 мегадалтона. Поради това че реципиентните клетки съществуват и се размножават нормално без присъствието на плазмиди, лесно могат да се определят свойствата, които плазмидите придават на клетките. Ето защо плазмидите са по-удобни за изследвания на структурата и функцията на генетичния материал in vitro, отколкото целите мутантни хромозоми на бактериите. Това подпомага генетичния анализ при бактериите. Някои плазмиди съдържат гени, осигуряващи пренасянето им в други клетки. Понякога междувидовите бариери не са преграда за проникването на плазмидите.
Транспозиция - преместването на малък фрагмент генетичен материал между нехомоложни участъци на хромозомата. Тя може да предизвика пълно или частично инхибиране на функцията на разположения до нея ген или гени, свързано с видима или летална мутация на тези гени. Понякога транспозицията може да доведе до активиране на близките гени. Транспозициите причиняват изменения на генотипа, поради което могат да се разглеждат и като мутации, и като рекомбинации. Те са обект на изследвания едва през последните години, поради което механизмите им са слабо проучени. Транспозиции са наблюдавани при прокариоти и еукариоти. Предполага се, че те имат почти универсално разпространение, но по-добре са изследвани при бактериите и при техните плазмиди и фаги. При еукариотните организми транспозициите са проучени при дрожди, дрозофила и царевица. Преместените при транспозицията фрагменти ДНК се наричат транспозиционни елементи и се означават с ТЕ. В зависимост от размерите им те се делят на малки, включващи 1-2 гена, наречени инсерционни сегменти (ИС-елементи), и големи транспозиционни елементи, наречени транспозони. Те имат централна част, ограничена от инвертирани (палиндромни) двойни подреждания при прокариотите и прави двойни подреждания при еукариотите.
Структура на ДНК при еукариотите
Изградената в процеса на еволюцията структура на ДНК на еукариотната клетка съществено се различава от структурата на ДНК на прокариотите. Прави впечатление огромната разлика в:
- Огромна дължина на молекулите ДНК, дължаща се на наличието на т.нар. мълчащи гени. При еукариотите съдържанието на ДНК на една клетка е средно с 2-3 степени по-високо, отколкото при прокариотите. Доказано е, че всяка хромозома съдържа една молекула ДНК. Увеличаването на количеството ДНК в еукариотните клетки вследствие на:
- Увеличението на броя на гените
- Наличието на повтарящи се подреждания на нуклеотидите и повишаването на дължината на използваните при транскрипцията цистрони съставлява около 103 пъти. Изчисленията показват, че ако при бактериите броят на структурните гени е (1-4).103, при висшите еукариотни организми той е (1-2).105 или дори малко повече. Една от причините за свръхвисокото количество ДНК в еукариотните клетки е наличието на т.нар. "мълчащи" гени, които служат на клетката при реализацията на рядко срещащи се генетични програми.
- Пакетиране на огромно количество ДНК в хромозомите чрез спирализация, което води до компактност на генома. Така дължината на генома в клетката не се различава съществено от дължината на пакетираната ДНК на прокариотите.
- Относителна нестабилност и пластичност - гени, с непостоянна локализация (скачащи гени). Проучени са преместванията на имуноглобулиновите гени през периода на индивидуалното развитие на мишки. Показател за пластичността на генома при еукариотните организми е амплификацията не само на рибозомните гени, но и на структурните гени, кодиращи някои белтъчини, в т.ч. и ензими. Амплификацията на гените се характеризира с образуването на многобройни копия (от 100 до 1000) от даден ген (например рРНК в ооцитите на различни видове организми) съобразно с потребностите на клетката. Но нестабилността на генома на еукариотите е твърде ограничена. Поради това, характерната структура на генома на даден вид организми се съхранява продължително време.
- амплификация на гените - образуване на многобройни копия на даден ген според потребностите на клетката;
- диференциация на ДНК по дължината на хромозомата - умерено повтарящи се и многократно повтарящи се подреждания на нуклеотидите.
- Уникално-повтарящи се подреждания на нуклеотидите - Те съставляват над 50% от генома на клетката. В тях са разположени структурните гени,които регулират биосинтеза на белтъчините. Те са подложени на активна транскрипция.
- Умерено-повтарящи се подреждания на нуклеотидите - Разпределят се между уникалните подреждания на нуклеотидите по дължината на молекулата на ДНК. Това са полимерните гени, които регулират унаследяването на количествените признаци при висшите организми. Те са значителна част от генома на еукариотите.
- Многократно-повтарящи се подреждания на нуклеотидите - Основно са локализирани в сателитната ДНК. Те се състоят от блокове къси участъци двойки нуклеотиди, които се повтарят многократно. Броят им в генома на еукариотите варира от милиони до десетки милиони. Разпределението им е видово-специфично. Всеки блок съдържа 6 нуклеотида (хексамер), а именно ТТТТТЦ/АААААГ. В резултат на това значителна част от цялото съдържание на ДНК в клетката се пада на сателитната ДНК. Тя е концентрирана неравномерно в определени места на хромозомата - в нейните сателити, в района на центромерите, теломерите и в малки количества в други части на хромозомите. Сателитната ДНК участва в изграждането на неактивния в генетично отношение конститутивен хетерохроматин. Върху молекулите сателитна ДНК не се осъществява транскрипция на никакви молекули РНК, защото тя не носи генетична информация. Разпределението й е видово специфично. Предполага се, че тя изгражда спейсерите, които разделят структурните и регулаторните гени, и играе роля в процесите на митозата и мейозата при прикрепването на хромозомите към нишките на делителното вретено или при разпознаването и свързването (конюгацията) на хомоложните хромозоми. Според някои автори сателитната ДНК защитава клетката от възникването на генни мутации. Освен сателитната ДНК в генома на еукариотите има малки количества многократно повтарящи се подреждания на нуклеотиди, които съдържат само двойки нуклеотиди АТ. Те кодират транскрипцията на къси участъци поли А-вериги, изградени само от нуклеотиди с аденин, с дължина 100-200 нуклеотида. Тези поли А-вериги се прикрепват към 3' - края на голяма част от молекулите на иРНК след тяхната транскрипция (т.нар. "опашка"), преминават заедно с тях от ядрото в цитоплазмата, но не се използват за транслация.
Основни генетични функции на ДНК
- Автокаталитична - репликация.
- Хетерокаталитична - транскрипция и транслация.
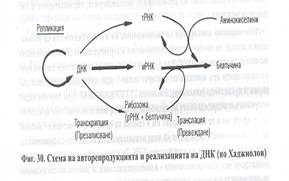
- Наследствената информация се предава от ДНК към белтъчините. Потокът на информацията е ДНК->РНК->белтъчини.