

Ключови думи:
химичен състав на клетката, биогенни елементи, гликозиди, естери, киселинни амиди (пептиди), киселинни анхидриди, структурна организация на клетката, прокариотна клетка, еукариотна клетка, клетъчни органели, ядро, цитоплазма, митохондрии, ендоплазмен ретикулум, апарат на Голджи, рибозоми.
Цели:
- Да се изясни химичния състав на клетката.
- Да се разгледат основните производни органични съединения и техните особености.
- Да научат строежа на прокариотната клетка.
- Да научат основните белези на структурата на еукариотните клетки.
- Да получат знания за клетъчните органели и техните функции.
Въведение
Клетката е най-малката структурна и функционална единица на живота, способна да се самовъзпроизвежда. Всички клетки съдържат множество по- малки структурни единици, наречени органели, които изпълняват специфични функции. Органелите са обкръжени с течна цитоплазма, а като цяло клетката е отделена от външната среда с липиднобелтъчна обвивка, наречена клетъчна мембрана. Цитоплазмената мембрана е жизнената преграда, която отделя вътрешността на клетката от външната среда. Вътре в клетката се намират цитоплазмата с клетъчните структури и вещества, които правят възможно нейното функциониране. Ключова структура е ядрото или нуклеоида, където се съдържа необходимата информация за репродукцията на клетката. В цитоплазмата са разположени механизми за клетъчния растеж, функциониране и размножаване.
Съществуват два големи класа клетки – прокариотни (доядрени) и еукариотни (същинско ядрени). Както показват имената им, разграничителен белег е наличието или липсата на оформено ядро, но всъщност разликите са по-дълбоки. Клетките в древните пластове са прокариотни, за което се съди по малките им размери и простото им устройство. Прокариотните са най-просто устроените малки клетки. Приема се, че тяхната възраст възлиза над 3,5 милиарда години. Еукариотните организми се появяват около милиард години след прокариотите.
1.1. Химичен състав на клетката
По химична природа веществата, които влизат в състава на клетката се разделят на две групи:
- неорганични компоненти: вода, органогенни елементи и минерални елементи;
- органични компоненти: белтъци, нуклеинови киселини, липиди, въглехидрати (полизахариди), алкохоли, витамини и др.
1.1.1. Неорганичен състав на клетката
Вода. Животът е възникнал във вода. Всички форми на живот на Земята не могат да живеят без вода. Водата участва в създаването на вътрешната среда на клетките на растения и животни. Свободната вода служи като разтворител на веществата в цитоплазмата на клетките. В нея се разтварят различни соли, които лесно се дисоциират на йони, а също и някои органични съединения. Тя изпълнява ролята на универсална дисперсна среда в клетката. При намаляване на съдържанието й до критично ниво, живите организми преминават в състояние на анабиоза.
Електронната й структура, физичните и химичните й свойства я правят пригодна за изпълнение на биологичната й роля. Молекулата на водата представлява леко "изкривен" тетраедър, в центъра на който е разположен кислородният атом. Двете му връзки с водорода са насочени към два от върховете на тетраедъра, докато несподелените електрони от двата sp3- хибридизирани орбитала заемат останалите два върха. Ъгълът между двата водородни атома (105o) е малко по-малък от съответния ъгъл при един правилен тетраедър (109,5o). Тази "дребна" разлика "изкривява" тетраедъра и е причина за повече от особените физико-химични свойства на водата, а оттам и за биологичните и функции. Електронният облак около кислородния атом e концентриран в област противоположна на водородните атоми. Така водната молекула придобива диполен характер.
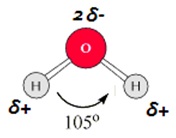
Фигура 1. Диполен характер на водната молекула - тетраедрична структура.
Всички вещества, които постъпват в клетката проникват с вода и само с вода се отделят продуктите от обмяната на веществата. Водата и йоните, получени от дисоциацията й са важни фактори, които определят структурата и функциите на биополимерите в клетката, в това число и на белтъците и нуклеиновите киселини. Водата може да промени пропускливостта на клетъчните мембрани, тъй като в резултат на различните и превръщания е възможно да настъпят изменения на съставящите ги компоненти.
Водата може да образува водородни връзки, които имат изключително голямо значение за биологичните системи (фиг. 2). Известно е, че повечето молекули в клетката образуват устойчиви „нежни" връзки само с неголям брой молекулни типове. Това частично се обяснява с водното обкръжение на молекулите в биологичните системи. Образуването на връзки между каквито и да са молекули в клетката зависи не само от това дали тези молекули могат да реагират помежду си, но също и от количеството на водородните връзки във водния разтвор.
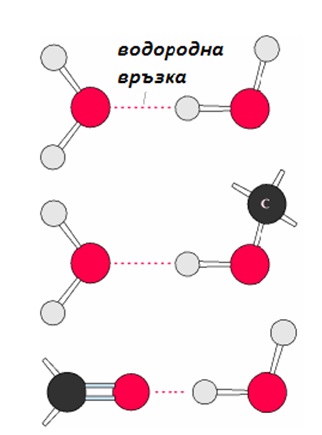
Фигура 2. Водородни връзки между диполите на водата
В биологичните системи водата изпълнява следните общи функции:
- Обединява всички части на организма;
- Разтворител и среда за извършване на биохимични реакции;
- Участва в създаването на порядък на структурите в клетките;
- Метаболит и непосредствен компонент на биохимичните процеси (при фотосинтезата, водата е донор на е-, необходима е за хидролизните и синтетичните процеси);
- Изпълнява функции в преносно-мембранните процеси;
- Главен компонент на транспортните системи на висшите организми;
- Терморегулиращ фактор, запазва тъканите от резки температурни колебания, благодарение на високата си топлоемкост;
- Добър “амортисьор” при механични въздействия върху организма.
Органогенни елементи - основни органогенни елементи са С, N, Н и О, които заемат от 86 до 98% от сухото вещество на клетките.
Въглерод - от 34 до 62% от сухото вещество на клетките, влиза в състава на повечето органични съединения и обуславя функциите им. Въглеродните атоми образуват помежду си прости (единични), двойни и тройни връзки, богати на енергия. Електронната му структура, неговите физични и химични свойства обуславят голямото разнообразие на органични съединения с широк диапазон на свойства. Въглеродът образува комплекси с голяма молекулна маса, които образуват колоидни разтвори, необходими за цитоплазмата и за построяване на фината и структура.
Азот - заема от 7 до 14% от сухото вещество на клетките, предимно в редуцирана форма (-NН2, -NН-, =N-) и придава реактивоспособност на органичните вещества, които го съдържат. Това се дължи на способността му трудно да влиза в химични съединения, но лесно да излиза от тях.
Кислород - заема около 30% и водород - от 5 до 8% от сухото вещество на клетките. Тези два елемента (единият окислител, а другият редуктор) вървят винаги заедно и влизат в разнообразни реакции с намиращите се около тях вещества.
Минерални елементи. Минералните елементи остават в пепелния остатък и заемат от 2 до 14% от сухото вещество на клетките. Това са: Р, Fе, S, Мg, Nа, К, Zn, Мn, Сu, Со и много други. Наличието на едни или други йони в клетките се обуславя от способността им да се пренасят от средата в клетката.
Една част от минералните елементи е необходимо да се доставят на човека с храната в по-големи количества над 100 mg ежедневно. Условно се наричат "макроминерали". Към макроминералите принадлежат шест елементи: четири метала - натрий, калий, калций и магнезий и два металоиди (неметали) - фосфор и хлор. Те се намират в организма обикновено в разтворено и дисоциирано състояние като катиони или аниони. Други са необходими в количества под 100 mg на ден, прието е да се наричат "микроминерали". Към микроминералите в човешкия организъм могат да се посочат: желязо, манган, мед, кобалт, цинк, селен, хром, молибден (катиони) и йод и флуор (аниони). Други елементи не са полезни за човека и дори някои от тях имат токсичен ефект.
Клетките съдържат значителни количества фосфор, който заема от 10 до 45% от пепелния остатък. Той се намира под формата на фосфорна киселина, свързана чрез естерни връзки с други атоми, фосфорът влиза в състава на клетъчните фосфолипиди, ДНК, РНК, нуклеопротеидите, редица ензими и др. Особено място сред елементите заема сярата, която рядко присъства в клетките във вид на минерални съединения. Обикновено тя влиза в състава на аминокиселините метионин, цистин и цистеин или е включена в някои вторични метаболити в сяросъдържащите хетероциклични съединения, а също и витамини (тиамин и биотин). Желязото е важен елемент за изграждането на дихателните ензими, цитохромите, цитохром- оксидазата, пероксидазата, каталазата.
Минералните елементи са с различно физиологично значение. Те стабилизират физико-химичното състояние на колоидите в цитоплазмата. Металните йони имат най-важно значение за функционирането на биологичните системи, свързани са с ензимните реакции. Те влизат в състава на редица ензими и по този начин участват в ензимни реакции. Магнезиевите йони (Мg+2) са необходими за съхраняването на устойчивите структури на рибозомите. Йоните на кобалта участват в състава на витамин В12. Калият е необходим за изграждането на някои повърхностни структури на микробните клетки. Той взема участие в набъбването на клетъчните белтъци. Йоните на някои минерални елементи се съдържат в клетките в много малки количества (т.нар. микроелементи).
1.1.2. Органични молекули изграждащи клетката
Главните химични компоненти на клетките са структури, наречени макромолекули. Основни биологични макромолекули са нуклеиновите киселини, белтъците, полизахаридите и липидите. Една част от тях са хомобиополимери (изградени чрез свързването на еднотипни, в някои случаи идентични основни структури), а другите хетеробиополимери, когато градивните единици при полимерите са еднотипни, но не идентични. Всеки един от тези полимери е изграден от характерно свързани мономери.
Макромолекулите на клетките се разделят на информационни и неинформационни. Информационни са нуклеиновите киселини и белтъците. В тях последователностите на мономерните единици са строго специфични, носят биологична информация и определят процесите на тази информация. Информационните молекули съдържат сравнително малък брой различни мономерни единици, при които важно значение имат техните последователности.
Полизахаридите и липидите не са информационни макромолекули, защото последователностите на техните мономери са във висока степен повтарящи се и не са функционално важни.
Основни органични структури. Органични съединения, които не подлежат на хидролиза, т.е. не притежават ковалентни връзки, които да се разкъсват при внасяне на вода – хидролиза, се наричат основни органични структури. Тези връзки могат да се разкъсват и чрез внасяне на фосфорна киселина, процес известен като фосфоролиза. В клетките не постъпват от околната среда производни органични структури, т.е. молекули, които биха могли да се подложат на хидролиза или фосфоролиза.
Основните органични структури (т.нар. мономери), са органични съединения, които съдържат до около 30 въглеродни атома и са групирани в класове в зависимост от техните химични особености. Има четири класа мономери, важни за клетката:
- монозахариди – мономери на полизахаридите
- нуклеотиди - базови единици в нуклеиновите киселини - ДНК и РНК;
- аминокиселини - мономери на белтъците;
- мастни киселини - мономерни единици на липидите.
Въглехидрати. Въглехидратите изграждат до 80% от сухото вещество на растенията: целулоза, нишесте, глюкоза, захароза, пектин и др. Висшите животни използват глюкоза, гликоген (резервна храна); рибозата и 2- дезоксирибозата се съдържат в нуклеиновите киселини. Наричат се още захари или захариди. Въглехидрати (англ. carbohydrates) буквално означава “хидрати на въглерода”, например глюкозата има молекулна формула (СН2О)6 = С6Н12О6.
Монозахариди (монози, захари). Това са полихидроксимоноалдехиди и полихидроксимонокетони. Според броя на въглеродните си атоми биват триози (с три въглеродни атома), тетрози (с четири), пентози (с пет), хексози (с шест), хептози (със седем) атоми. Най-застъпени в природата са пентозите и хексозите.
Природните монозахариди са разпространени само в оптичноактивна форма и предимно един от възможните енантиомери се среща в природата. Енантиомерите (оптичноактивен изомер) са неразличими физически и химически, но се отнасят различно, когато лъч на плоско-поляризована светлина се пропусне през разтвор на оптично активен изомер. Наличието на асиметрични атоми обуславя свойството оптична активност - способност да въртят равнината на поляризована светлина. Посоката и ъгълът на въртене се измерват с уред, наречен полярометър. Изомерите могат да бъдат дясновъртящи (+) или лявовъртящи (-). Няма ясна зависимост между структурата на молекулата и посоката на въртене. Така че дадено съединение може да бъде D(+), D(-), L(+) или L(-).
Смес от равни количества от D и L форми няма оптична активност, тъй като активностите на двата изомера се неутрализират. Такава смес се нарича рацемична. Обикновено ензимите различават и действат само на единия от двата изомера.

Епимери се наричат двойка диастереоизомери, които се отличават помежду си по конфигурацията на само един от хиралните центрове (наричани понякога още “стереогенни центрове”).
Нуклеинови киселини. Нуклеиновите киселини са биополимери - дезоксирибонуклеиновите киселини (ДНК) и рибонуклеиновите киселини (РНК). Дезоксирибонуклеиновите киселини (ДНК) се съдържат в ядрата на всички еукариотни клетки, съхраняват и предават генетичната информация от поколение на поколение. Рибонуклеиновите киселини (РНК) преписват и пренасят информацията от ДНК, спомагат за синтеза на белтъците. При меко ензимно разграждане се получават съответните мономери, наречени нуклеотиди.
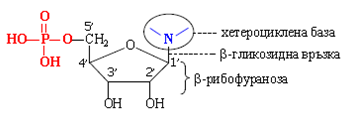
Нуклеотиди. Нуклеотидът се състои от три фрагмента:
- Пиримидинова или пуринова хетероциклена база, свързана с пентозния остатък чрез β-N-гликозидна връзка. Пиримидиновата база се свързва при N-1, а пуриновата – при N-9.
- Пентозен остатък: D-рибофураноза (РНК) или 2´-дезокси-D- рибофураноза (ДНК). Въглеродните атоми на пентозата се номерират с „прим” (1´, 2´ и т. н.).
- Остатъци на фосфорната киселина – естерифицирани са хидроксилни групи на 3´- и/или на 5´ -място.
Обща формула на нуклеотид:
Белтъци. Както прокариотните, така и еукариотните клетки съдържат значителни количества белтъци. Най – голям дял сред органичните съставки имат белтъците, достига 65 - 70% и повече.
Белтъците имат първостепенно значение при формирането на структурата на клетките и обезпечават проявата на жизнените функции. Белтъците са молекулите, с помощта на които се реализира генетичната информация. Някои от белтъците служат като резервни хранителни вещества, а други, каквито са пермеазите извършват транспортиране на хранителни вещества и метаболитни продукти в клетката или вън от нея. Голяма част от белтъците като ензими проявяват специфични каталитични функции. Белтъците са структурни елементи на клетката, разпределени в цитоплазмата, ядрото, мембранните структури и др. В клетките важно значение имат съединенията на белтъците с други полимери, каквито са липопротеидите, глюкопротеидите, нуклеопротеидите. В ядрото се съдържат нуклепротеиди, известни като хистони.
Аминокиселини. Аминокиселините по строеж са органични карбонови киселини, при които като минимум, един атом водород е заместен с аминогрупа. Аминокиселините са структурни мономери на белтъчните молекули, но необходимостта от изучаването им не се дължи само на тази функция. Някои от тях се явяват източници за образуване на навромедиатори в ЦНС (хистамин, серотонин, гама-аминомаслена киселина, допамин, норадреналин), други сами се явяват невромедиатори (глицин, глутаминова киселина). Аминокиселини участват в синтеза на пуринови и пиримидинови бази, които влизат в състава на нуклеиновите киселини, и се използват за синтез на низкомолекулни биологично важни съединения (креатин, карнитин, карнозин, ансерин и др.). Аминокиселината тирозин изцяло влиза в състава на хормоните на щитовидната жлеза.
Липиди. Липиди са отрити във всички живи молекули и играят важна роля в поддържането на живота. За разлика от въглехидратите и протеините, липидите са високо полиморфни и структурно е трудно да се определят. В резултат на това, те се дефинират съгласно тяхната основна характеристика като неразтворими във вода органични съединения открити в биологичните системи. Липидите могат да бъдат или амфипатични (имащи полярни и неполярни качества) или хидрофобни (неполярни).
Независимо, че липидната структура е относително сложна, съществуват няколко общи структурни зависимости (фиг.3). Най-простите липиди са масни киселини с обща формула R-COOH, където R е въглеводородна опашка (верига, радикал). Мастните киселини могат да бъдат съставни части на по- сложни липидни комплекси, такива като триацилглицероли, глицерофосфолипиди, свинголипиди, восъци и ейкозаноиди. От друга страна, съществуват структурно различни липиди, такива като стероидите и липидните витамини, които произлизат от пет въглеродната молекула на изопрена. Липидите съдържащи фосфатна част се наричат фосфолипиди, липиди с въглехидратна част се наричат гликолипиди и липиди произлизащи от изопрена се наричат полипренилни съединения (или изопреноиди).
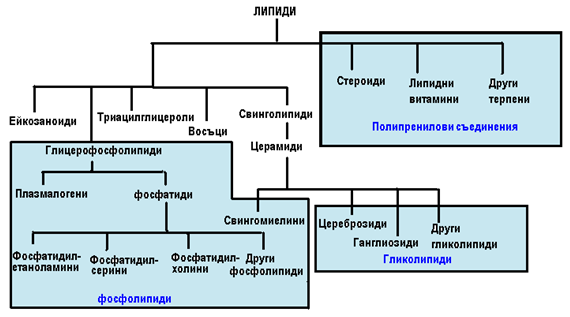
Фигура 3. Основни видове липиди.
Мастни киселини (Карбоксилови киселини). Обикновено това са алифатни монокарбоксилови киселини. Те са един от двата (наред с монозахаридите) основни източници на енергия за организмите. Голямо значение имат и като съставки на сложните (комплексни) липиди (фиг.3): триацилглицероли, фосфолипиди, холестеролови естери и др. Мастните киселини са предимно с четен брой въглеродни атоми, и то най-вече такива с по-голям брой от 14 въглеродни атоми, наречени висши мастни киселини.
Някои от монокарбоксиловите киселини съдържат двойни връзки - ненаситени мастни киселини. Наричат се общо “енови” киселини: моноенови с една, диенови с две, триенови с три, тетраенови с четири и т.н. двойни връзки.
1.1.3. Свързване на основните органични структури в производни
В природата броят на органичните съединения с производни структури е значително по-голям. Основните органични структури се свързват помежду си по два принципно различни механизма, известни в органичната химия като полимеризация и кондензация.
Полимеризацията се дължи най-често на разкъсване на двойна връзка, т.е. изисква наличие на ненаситени съединения.
Кондензацията е процес, при който основните органични структури се свързват чрез отделяне на молекула вода (или отделяне на фосфорна киселина) между реагиращите помежду си функционални групи. Съответно на това се получават и четири типа производни полимери (респ. поликондензати): гликозиди; естери; киселинни амиди (респ. пептиди) и киселинни анхидриди. В живата природа са познати четири начина за свързванe чрез кондензация, при които се получават и четири типа връзки:
Y гликозидна – получава се при обезводняване на гликозидната (полуацеталната) хидроксилна група на монозахарид с всякаква друга хидроксилна, тиолова, или циклична аминогрупа. Въглехидратите при образуването на гликозидни производни реагират със своите циклични (полуацетални) структури. Най-застъпени в природата са гликозидните връзки с хидроксилни групи. Това са така наречените кислород-гликозиди. Кислород-гликозидни са връзките при всички олиго- и полизахариди.
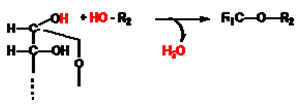
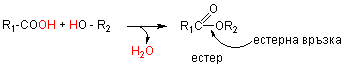
Y естерна - получава се от обезводняване на алкохолна група с различни киселини: карбоксилови, фосфорна, сярна и др..; Естерите са много застъпена в природата кондензати. Естери с карбоксилови киселини са триацилглицеролите, восъците и етеричните масла. Естерна връзка има и при стероидите - например естерифициран с мастна киселина холестерол.
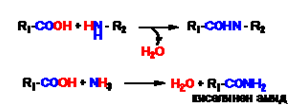
Y киселинноамидна (в някои случаи наричана пептидна) - Получава се при обезводняване на карбоксилна и амино-групи или амоняк. Когато киселинноамидната връзка се образува между алфа-амино и алфа- карбоксилната група на две алфа-аминокиселини, тя се нарича пептидна връзка.
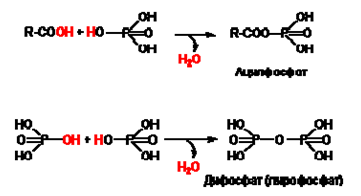
Y киселинноанхидридна - получава се при обезводняване на две химични групи с “киселинен" характер. Киселинно-анхидридните връзки са твърде нестабилни. Съединенията, които ги съдържат, се наричат макроергични съединения.
1.2. Структура на клетката – прокариотна и еукариотна
Съществува голямо разнообразие от клетки по отношение на форма, размери и функционална специализация, но независимо от това изумителното разнообразие всички те поразително си приличат по основните структури, които ги изграждат. Всяка клетка притежава клетъчна (плазмена) мембрана, ядро и цитоплазма.
1.2.1. Прокариотни клетки
Най-характерната особеност на прокариотите е отсъствието на обособено ядро с ядрена мембрана и ядрена плазма. При тях ядрената материя представлява огромна молекула на ДНК, която е разположена в цитоплазмата и в определена точка е закрепена за цитоплазмената мембрана. ДНК не е свързана с белтъци (хистони) и представлява своеобразен силно нагънат пръстен, като формира т.нар. ядрена област и се нарича нуклеоид (фиг. 4). Наличието само на една хромозома при прокариотите е предпоставка да се отнасят към хаплоидните клетки.
Прокариотната клетка съдържа пет основни структурни компонента (фиг.4):
- Нуклеоид (ДНК);
- Рибозоми;
- Клетъчна мембрана;
- Клетъчна стена;
- Клетъчни повърхностни структури.
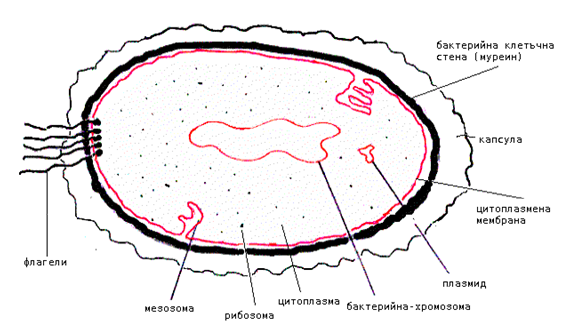
Фигура4. Структура на прокариотната клетка
1.2.2. Еукариотни клетки
Характерна особеност на еукариотните клетки са по-големите им размери, наличието на диференцирано ядро с ядрена мембрана и плазма. ДНК, която изгражда ядреното вещество, е свързана с белтъци и е накъсана на отделни елементи, наречени хромозоми. Еукариотните клетки съдържат специфични обособени структури, като митохондрии, ендоплазмен ретикулум, апарат на Голджи, вакуоли, пероксизоми и др. (фиг.5а – раститерна клетка; 5б – животинска клетка).
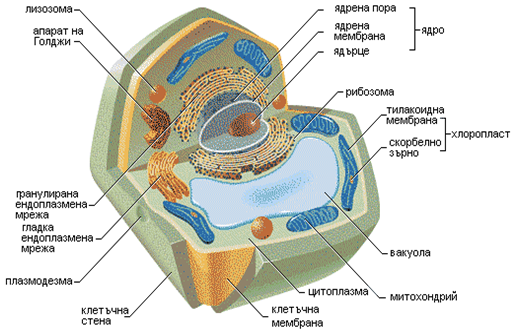
Фигура 5а. Структура на растителна клетка
Всички многоклетъчни организми са изградени от еукариотни клетки, за които е характерно делене чрез митоза и наличие на двоен набор от хромозоми, произхождащи от родителските им клетки. За това носят названието диплоидни клетки.
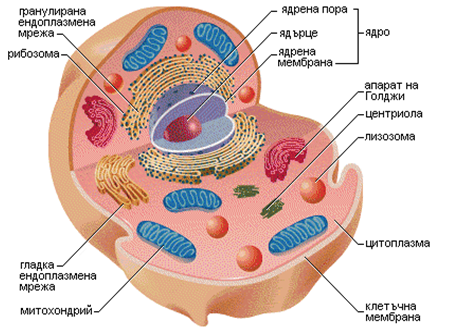
Фигура 5б. Структура на животинска клетка
1.3. Клетъчни органели – структура и функции
1.3.1. Ядро – структура и функции
Ядрото е най-сложната структура на клетката. В него е разположена ДНК, чиято основна функция е да запазва и предава в наследство записаната в нея генетична информация на клетката. В ядрото ДНК се намира свързана с белтъци и други молекули под формата на хроматин, който представлява тънки нишки. Преди деление на клетката хроматинът се групира в добре оформени образувания - хромозоми. През останалото време той е разпръснат в безпорядък. Прокариотите нямат морфологично оформено ядро (ограничено с ядрена мембрана), но те също съдържат ДНК, която се локализира в определено място на клетката и се означава като нуклеоид.
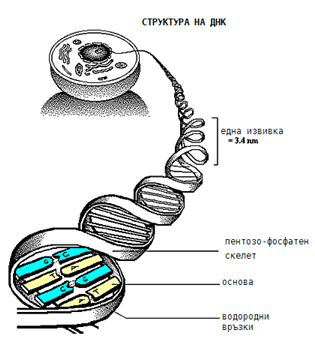
Фигура 6. Структура на ДНК при еукариотите
Ядрото на еукариотните клетки съдържа едно или две морфологично обособени ядърца. Съдържанието на клетъчното ядро (нуклеоплазма) е отделено от цитоплазмата с ядрена обвивка. Тя се състои от две близко разположени мембрани, разделени с тясно пространство (20-40 nm). Голяма част от външната ядрена мембрана е свързана с ендоплазмения ретикулум. В определени участъци на ядрената обвивка двете мембрани се сливат една с друга и се образуват ядрени пори с вътрешен диаметър около 80 nm. През тях преминават лесно високомолекулни вещества от ядрото в цитоплазмата и обратно. Това е особено важно за синтезираните вътре РНК, които след известни промени (познати като процесинг) преминават в цитоплазмата и се използват при синтеза на белтък.
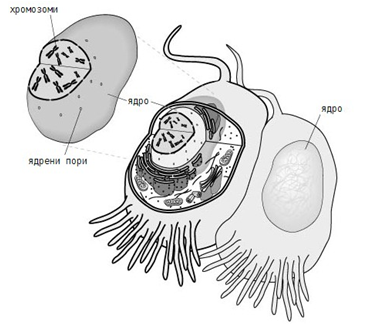
Фигура 7. Ядро и ядрена обвивка
1.3.2. Цитоплазма – състав и строеж
Цитоплазмата се състои от:
- Основно вещество (цитозол);
- Клетъчни органели;
- Клетъчни включения;
- Цитоскелет;
Основната част от обема на клетката се заема от цитоплазмата. Тя изгражда вътрешното пространство на всяка клетка и представлява необикновено сложна смес от големи и малки молекули. Съдържа около 70% вода и 15-20 % белтък (по тегло). В нея протичат голяма част от ензимно катализираните реакции на метаболизма. Чрез тях се освобождава химическа енергия, необходима за цялостната жизнена дейност на клетката.
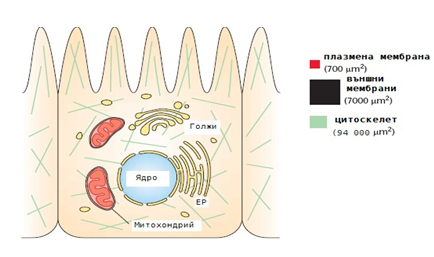
Фигура 8. Основни компоненти на архитектурата на еукариотната клетка.
Цитоплазмата е високо организирана структура, в която се откриват множество разнообразни органели и включения. В прокариотните клетки те са представени основно от рибозомите и различните въглехидратни, липидни и др. включения. Еукариотните клетки имат значително по-сложен вътреклетъчен строеж. Освен рибозоми те притежават още ендоплазмен ретикулум, митохондрии, лизозоми, апарат на Голджи, пероксизоми, хлоропласти и вакуоли (само в растителните клетки) и др.
Остатъкът, който остава след отстраняването на клетъчните органели се означава като цитозол. Той заема около 55 % от обема на клетката» съдържа стотици ензими, които катализират реакциите на гликолизата и глюконеогенезата, биосинтезата на захари, мастни киселини, нуклеотиди и аминокиселини. Тук са разположени и нискомолекулните тРНК, които пренасят аминокиселините към рибозомите. В цитозола е установено присъствието на система от микротубули и микрофиламенти, които формират цитоскелета. Той е изграден от белтъци и чрез него се поддържа формата на клетката, подпомага се осъществяването на много ензимни реакции и се участва в клетъчното движение. Присъствието в цитозола на 15-20 % белтък го превръща по-скоро във високоорганизирана желеобразна маса, отколкото просто разтвор на ензимни молекули. Заедно със строителните блокчета на биополимерите (монозахариди, аминокиселини, нуклеотиди) в цитозола се намират и стотици малки молекули на органични съединения, промеждутъчни продукти на метаболизма. Освен тях се установява присъствието на различни кофактори на ензимите, на главните компоненти от системата за пренос на енергия — АТР и АDP а също така и на голям брой неорганични йони (К+, М++, Са++, Сl-, НСО3-и др.).
1.3.3. Рибозоми – структура и биологична функция
Рибозомите са немембранни клетъчни клетъчни органели, компактни частици изградени от рибозомна РНК (рРНК) и белтъци, чиято функция е свързана със синтеза на белтъци. Рибозомите се срещат, както в прокариотни, така и в еукариотни клетки. Всяка рибозома се състои от две субединици - голяма и малка, свързани помежду си с магнезиеви йони, които обратимо се дисоциират след завършване синтезата на даден полипептид. Рибозомите заедно с гранулирания ендоплазмен ретикулум и апарата на Голджи участват в синтезата на клетъчни протеини. В тях се осъществява процесът транслация – т.е. превеждане на генетичната информация от полинуклеотидния код на нуклеиновите киселини в аминокиселинния код на белтъците. Различават се два вида рибозоми - свързани и свободни. Свързаните са закрепени с голямата си субединица по-външната мембранна повърхност на гранулирания ендоплазмен ретикулум, те продуцират секреторни белтъци - Ig, хормони и други. Свободните рибозоми са единични или на групи от 5-9 (т. нар. полизоми). Свободните рибозоми продуцират белтъци за самата клетка.
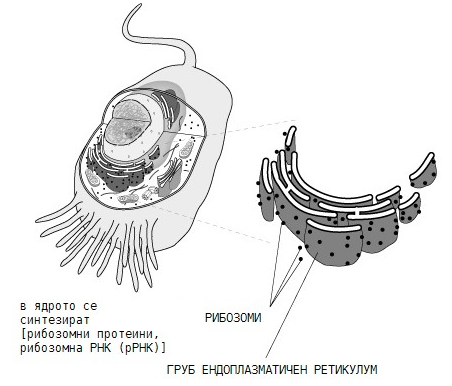
Фигура 9. Синтез и локализация на рибозомите
Рибозомите на еукариотите и прокариотите имат приблизително еднаква структура и функции. Различават се по своите размери и състав. Прокариотната рибозома е с коефициент на седиментация 70S. Голямата й субчастица (50S) е изградена от две рРНК (23S и 5S) и 34 белтъка, а малката (30S) - от една рРНК (16S) и 21 белтъка. Еукариотната рибозома е значително по-голяма от прокариотната и има коефициент на седиментация 80S. Нейната голяма субчастица (60S) включва в състава си три рРНК (28S; 5,8S и 5S) и 40 белтъка, а малката субчастица (40S) е изградена само от една рРНК (18S) и 33 белтъка. При този състав на рибозомите съотношението между РНК и белтъка в еукариотите е около 1, докато прокариотите са по-богати на РНК и съотношението е около 2. По-голяма част от рибозомите в еукариотните клетки са прикрепени към външната страна на мембраните на ендоплазмения ретикулум, но в цитоплазмата има и свободни рибозоми, в което състояние те се намират във всички прокариоти. Освен в цитоплазмата, рибозоми има и в ядрото на клетката, митохондриите и хлоропластите.
1.3.4. Митохондрии – структура и функции
Митохондриите са вътреклетъчни органели, присъщи само на еукариотите. Те са едни от най-подробно проучените в морфологично, биохимично и функционално отношение структури. Размерите им (0,5-2 μm) позволяват да бъдат наблюдавани дори с обикновен светлинен микроскоп. Митохондриите са подвижни органели и тяхното движение е свързано с микротубулите на цитоскелета. Митохондриите изпълняват в клетките ролята на “силовите станции”, в които се генерира преобладаващата част от необходимата енергия, под формата на макроергични връзки.
Всяка митохондрия има две високоспециализирани мембрани, които играят ключова роля в нейната активност. Външната мембрана е гладка, а вътрешната се сгъва навътре, образувайки напречни прегради - кристи. В резултат на това нагъване площта на вътрешната мембрана нараства многократно и повишава нейните възможности за синтез на АТР. Броят на кристите за различните видове клетки е различен. В сърдечните клетки техният брой е три пъти по-голям от тези в черния дроб.
В състава на вътрешната мембраната са включени:
а) белтъците, катализиращи окислителните реакции в дихателната верига;
б) ензимният комплекс, който синтезира АТР — АТР-синтезата;
в) специфични транспортни белтъци, осигуряващи преноса на метаболити в матрикса и извън него.
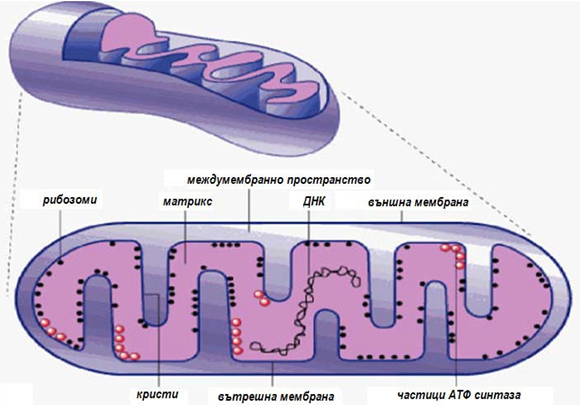
Фигура 10. Структура на митохондрий
Пространството в митохондрия, заградено от вътрешната мембрана е запълнено с желеподобно вещество - матрикс. То съдържа високо концентрирана смес от стотици ензими в т.ч. и ензимите, необходими за окисление на пирувата, на висшите мастни киселини и за провеждане на цикъла на лимонената киселина. В матрикса са разположени още специфични рибозоми, няколко идентични копия на митохондриална ДНК, тРНК и различни ензими, участващи в биосинтеза на специфични белтъци на вътрешната мембрана. Между двете мембрани се образува тясно междумембранно пространство, което е бедно на белтъци и съдържа няколко ензима (аденилаткиназа и др.), които използват излизащите от матрикса молекули АТР за фосфорилиране на други нуклеотиди. Тук постъпват също и различни нискомолекулни съединения (с молекулна маса до 10 000) — кофактори, субстрати на биологичното окисление и нискомолекулни белтъци. Те преминават сравнително лесно, тъй като външната мембрана съдържа много молекули транспортни белтъци, образуващи широки хидрофилни канали в двойния липиден слой. В нейния състав влизат също ензими, които превръщат липидите в реактивоспособни интермедианти, участващи в метаболитните процеси в матрикса.
1.3.5. Ендоплазмен ретикулум (мрежа)
Ендоплазмен ретикулум (мрежа) има само в еукариотните клетки. Неговата мембрана заема над половината от общото количество клетъчни мембрани. Тя образува многобройни дипли и разклонения, които обаче изграждат една непрекъсната затворена повърхност, наподобяваща слоеста торбичка. Вътрешното пространство на ендоплазмения ретикулум (ЕР), наречено цистерни заема над 10% от общия обем на клетката и се отделя от цитозола с единична мембрана. Тя образува с външната ядрена мембрана непрекъсната структура.
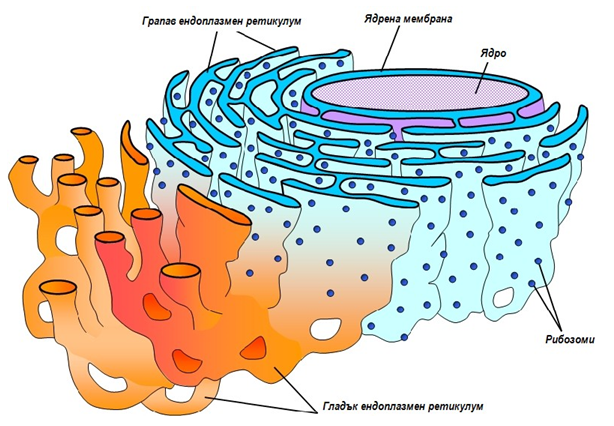
Фигура 11. Грапав и гладък ендоплазмен ретикулум
Съществуват два типа ендоплазмен ретикулум — грапав и гладък. Външната повърхност на мембраната на грапавия ЕР е осеяна с рибозоми, докато в гладкия ЕР рибозоми няма. Двата вида ЕР се различават и по форма — грапавият ЕР обикновено изглежда като система от плоски слоеве, които са покрити с рибозоми, а гладкият ЕР се състои от мрежа тънки тръбички. Ендоплазменият ретикулум е специализиран в синтезата и транспорта на липиди и мембранни белтъци, които се използват за собствени нужди, за изграждане на другите клетъчни органели (апарат на Голджи, лизозоми, плазмена мембрана и др.) или се изнасят в извънклетъчното пространство. Върху рибозомите, свързани с мембраните на грапавия ЕР се синтезират разнообразни белтъци. Те се изтласкват през мембраната във вътрешността на цистерните, откъдето се разпределят по направление.
Основната роля на гладкия ЕР е свързана с метаболизма на липидите. Ензимите, които осъществяват синтеза на липидните компоненти на липопротеините са локализирани в мембраните на гладкия ЕР При някои клетки в тях се разполагат и ензими, катализиращи реакциите на детоксикация. В резултат на това се обезвреждат лекарствени вещества, а също така и вредни метаболитни съединения.
В различните типове еукариотни клетки ЕР се различава по форма и изпълнява преимуществено определени функции. Например, в клетките на задстомашната жлеза и тези, секретиращи антитела преобладава грапавият ЕР, към повърхността на неговите мембрани са свързани над половината от всички рибозоми на клетката. Цистерните на ЕР в тези клетки са силно разширени, за да поемат всички белтъци, които преимуществено се синтезират в тях.
1.3.6. Апарат на Голджи
Апарат на Голджи обикновено е разложен близо до клетъчното ядро. В нормални условия той представлява група плоски, сплескани мембранни торбички, наричани цистерни. Всяка група в тях образува структура, наподобяваща купчина чинии, наричани още телца на Голджи. Една купчина обикновено съдържа 6 цистерни с диаметър около 1μm, но в някои низши еукариоти техният брой, може да достигне до 30. В различните клетки броят на телцата на Голджи варира в широки граници от едно до няколко стотици.
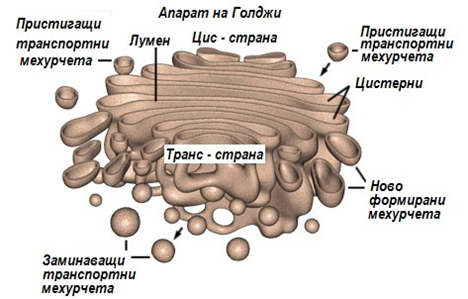
Фигура 12. Структура на Апарата на Голджи
Телцата на Голджи винаги са обкръжени с асоциирани към тях голям брой малки мехурчета (диаметър около 50 nm), повече или по-малко сферични и ограничени с мембрана. Счита се, че те транспортират материал до апарата на Голджи и извън него, към останалите клетъчни компоненти. Апаратът на Голджи има три дефинирани области — цис, междинна и транс. Цис-областта е близо до ЕР и се свързва с транспортните мехурчета, идващи от него. Тяхното съдържание се пренася през междината и достига до транс областта, откъдето то се отправя към определени места в клетката и извън нея. Апаратът на Голджи вероятно е главният "регулировчик" на клетката. Той играе ключова роля в сортирането на клетъчните белтъци и мембранните съставни части и отправянето им към техните точни предназначения. Голям брой от ензимите в апарата на Голджи ковалентно модифицират преминаващите през него секреторни или мембранни белтъци и гликопротеини. В повечето случаи промените засягат олигозахаридната фракция или са свързани с прибавянето на определен "маркер" или "сигнал" (например, фосфатен остатък), чрез който се постига разпознаването и попадането им на точно определени места.
1.3.7. Лизозоми
Лизозомите са обкръжени с мембрана, която формира сферични торбички, съдържащи хидролази, чрез които се осъществява контролирано, вътреклетъчно разграждане на вече ненужни на клетката белтъци, полизахариди, липиди, нуклеинови киселини. Лизозомите, които имат различна размери (0,1-0,5 μm) и форма, съдържат протеази, нуклеази, глюкозидази, липази, фосфолипази, фосфатази и сулфатази. Техният рН оптимум е около 5. Досега са изолирани над 40 от тези ензими. В нормални условия цитоплазмата на клетката е защитена от тяхното действие, тъй като мембраната на тези органели е непроницаема за тях. Другият защитен механизъм е свързан с рН на клетката, което е около 7. При случайно разкъсване на лизозомната мембрана, киселите хидролази попадат в неутрална среда и почти не проявяват активност т.е. не могат да действат. Характерно за мембраната на лизозомите е, че тя позволява да я напускат лесно продуктите от разграждането на макромолекулите. Тези разградни метаболити могат да се отделят извън клетката или да се използват от нея, Освен това по повърхността на мембраната се намират особени белтъци- маркери, към които се насочват сливащите се с нея специфични транспорти мехурчета. Съществуват два основни класа лизозоми:
а) първични, които имат приблизително сферична форма и още не са срещали субстрат за разграждане;
б) вторични, които са по-големи, имат неправилна форма и съдържат субстрати и хидролитични ензими.
След смъртта на клетките лизозомите се разрушават и техните хидролази извършват автолиза на клетките.
1.3.8. Пероксизоми
Пероксизомите са малки, обкръжени с мембрана органели, които се намират в цитоплазмата на всички животински клетки и на малък брой растителни клетки. Дълго време те са смятани за лизозоми, тъй като морфологията на двете групи органели е твърде сходна. Едва в началото на 1960 г. се установява, че пероксизомите се различават много по състав на ензими и функции от лизозомите. Пероксизомите, подобно на митохондриите, са един от центровете за усвояване на кислорода в клетката. Те съдържат ензими, които окисляват мастни киселини, аминокиселини и други субстрати, при което се получава Н2О2 . За предпазване от вредното действие на водородния пероксид върху клетките, пероксизомите съдържат изобилно количество каталаза, която го разгражда. В прорастващите семена присъства специфична група пероксизоми, които превръщат резервните липиди в захари. Това става чрез серия от реакции, известни като глиоксалатен цикъл, поради което тези пероксизоми носят името глиоксизоми.
В клетките се намират и други по-общи или по-специализирани структури - въглехидратни включения (под формата на гранули от нишесте и гликоген, липидни включения (под формата на мастни капчици), вакуоли, миофибрили, неврофибрили и др.
Терминологичен речник (Ключови понятия ):
# прокариотна клетка - най-характерната особеност на прокариотите е отсъствието на обособено ядро с ядрена мембрана и ядрена плазма. При тях ядрената материя представлява огромна молекула на ДНК, която е разположена в цитоплазмата и в определена точка е закрепена за цитоплазмената мембрана. ДНК не е свързана с белтъци (хистони) и представлява своеобразен силно нагънат пръстен, като формира т.нар. ядрена област и се нарича нуклеоид.
# еукариотна клетка – характеризира се с по-големи размери, наличие на диференцирано ядро с ядрена мембрана и плазма. ДНК, която изгражда ядреното вещество, е свързана с белтъци и е накъсана на отделни елементи, наречени хромозоми. Еукариотните клетки съдържат специфични обособени структури, като митохондрии, ендоплазмен ретикулум, апарат на Голджи, вакуоли, пероксизоми и др.
# ядро - най-сложната структура на клетката, в него е разположена ДНК, чиято основна функция е да запазва и предава в наследство записаната в нея генетична информация на клетката.
# митохондрии - вътреклетъчни органели, присъщи само за еукариотите, изпълняват в клетките ролята на “силови станции”, в които се генерира преобладаващата част от необходимата енергия, под формата на макроергични връзки на АТФ.
# лизозоми - обкръжени с мембрана, която формира сферични торбички, съдържащи хидролази, чрез които се осъществява контролирано, вътреклетъчно разграждане на вече ненужни на клетката белтъци, полизахариди, липиди, нуклеинови киселини.
# пероксизоми - малки, обкръжени с мембрана органели, които се намират в цитоплазмата на всички животински клетки и на малък брой растителни клетки. Съдържат ензими, които окисляват мастни киселини, аминокиселини и други субстрати, при което се получава Н2О2. За предпазване от вредното действие на водородния пероксид върху клетките, пероксизомите съдържат изобилно количество каталаза, която го разгражда.
# рибозоми - компактни частици изградени от рибонуклеопротеини и функцията им е свързана със синтеза на белтъци. Всяка рибозома се състои от две субединици, голяма и малка субчастица, които обратимо се дисоциират след завършване синтезата на даден полипептид.
# апарат на Голджи - вероятно е главният "регулировчик" на клетката. Той играе ключова роля в сортирането на клетъчните белтъци и мембранните съставни части и отправянето им към техните точни предназначения.
# ендоплазмен ретикулум (мрежа) - само в еукариотните клетки, като заема над половината от общото количество клетъчни мембрани. Съществуват два типа ендоплазмен ретикулум — грапав и гладък. Върху рибозомите, свързани с мембраните на грапавия ЕР се синтезират разнообразни белтъци. Основната роля на гладкия ЕР е свързана с метаболизма на липидите.
Органогенни елементи - основните органогенни елементи са С, N, Н и О, които заемат от 86 до 98% от сухото вещество на клетките.
# Минерални елементи - остават в пепелния остатък и заемат от 2 до 14% от сухото вещество на клетките, това са: Р, Fе, S, Мg, Nа, К, Zn, Мn, Сu, Со и много други.

Проект „Развитие на център за електронни форми на дистанционно обучение в Тракийски университет“ BG051PO001-4.3.04-0026
