

Ключови думи: анаеробно и аеробно окисление на въглехидратите, гликолиза, пентозофосфатен път, глюконеогенеза
Цели:
1. Да се разгледа анаеробното и аеробно разграждане на въглехидратите.
2. Да се изясни химизма и ролята за метаболизма на гликолизата.
3. Да се изясни химизма и ролята на фотосинтезата.
4. Да се изясни химизма и ролята на глюконеогенезата.
5. Да се изясни химизма и ролята на пентозофосфатният път.
Въведение
Въглехидратите са полихидрокси алдехиди, кетони или субстанции, които при хидролиза се разграждат до такива мономери. Повечето, но не всички имат емпирична формула (CH2O)n. Някои съдържат азот, фосфор, или сяра. Въглехидратите са предпочитан източник на енергия за организмите. Основен субстрат за получаване на енергия в клетката е глюкозата. Окислението на глюкозата протича по два пътя:
Ø Окисление с получаване на енергия (гликолиза);
Ø Оксиление с образуване на пентози: рибози, рибулози, ксилулози.
Този път се нарича пентозофостфатен път (ПФП) и не е свързан с получаване на енергия.
Гликолизата е катаболитен път на анаеробно окисление на глюкозата, появила се преди повече от 3,5 млрд. години, свързана синтез на АТФ и завършва с образуване на пируват. При анаеробни условия, пирувата се редуцира до лактат, което е съпроводено с окисление на НАДН до НАД+. При аеробни условия, посредством окислително декарбоксилиране на пируват до ацетил-КоА, лигазно карбоксилиране на пируват до оксалацетат и редуктивно карбоксилиране на пируват до малат, може да се свърже с цитратния цикъл (ЦК).
Пентозофосфатният път (ПФП) обезпечава клетките с рибози и НАДФН. ПФП окислява глюкозата и при определени условия може напълно да я разгради до СО2 и Н2О, но се класифицира като анаболитен път, с основни функции:
Ø образуване на редуциран НАДФH, необходим за биосинтези;
Ø снабдяване с рибозо-5-фосфат за синтеза на нуклеинови киселини;
Ø образуване на субстрати за гликолизата и гликонеогенезата;
Този метаболитен път не се използва за образуване на енергия, тъй като е единствeн път за синтез на рибоза и основен път за натрупване на НАДФ.Н.
Синтезата на въглехидрати от неорганични вещества се осъществява от автотрофните организми, за сметка на енергията от слънцето, и е известна като процес наречен фотосинтеза. Фотосинтезата е биологичен процес, характерен за зелените растения и някои микроорганизми (цианобактерии), дефинира се като физико-химичен процес, при който фотосинтезиращите организми използват светлинната енергия за да извършват синтез на органични вещества от неорганични.
Синтезата на глюкоза от невъглехидратни предшественици (лактат, пируват, α-кетокиселини от цитратния цикъл) и α-кетокиселини от аминокиселините с изключение на левцин и лизин, в хетеротрофните организми, е метаболитен път известен като глюконеогенеза.
11.1. Анаеробно и аеробно разграждане на въглехидратите
Гликолизата е анаеробен, основен катаболитен път на разграждане на глюкоза в клетките, нарича се още фосфотриозен, или метаболитен път на Ембден–Майерхоф–Парнас (ЕМП). Названието «гликолиза» произхожда от гр. γλυκός, glykos — сладко и λύσης, lysis — разграждане.
Това е един от най-древните метаболитни процеси, известен почти при всички живи организми. Предполага се, че се е появила преди повече от 3,5 млрд. години. Представлява анаеробно окисление на глюкоза, съпроводено със синтез на АТФ и завършва с образуване на пируват.
В зависимост от това дали пирувата се редуцира до млечна киселина, при отсътвие на кислород (т.е. анаеробни условия), или ще се превърне в ацетил-SКоА в присъствие на кислород (т.е. аеробни условия), окислението на глюкозата се класифицира като анаеробно или аеробно съответно (фиг.156).
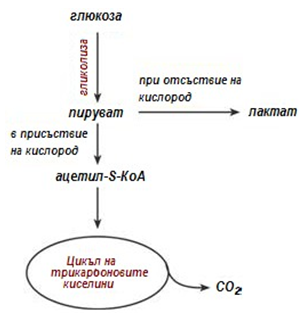
Фигура 156. Анаеробно и аеробно разграждане на въглехидратите
При аеробно окисление на глюкозата, пирувата се превръща в ацетил- SКоА от ПВК-дехидрогеназа и след това се включва в реакциите на цикъла на трикарбоновите киселини като се окислява до СО2. Сумарното уравнение на аеробното окисление на глюкозата, може да се представи като:
С6Н12О6 + 6 О2 + 38 АДФ + 38 Фнеорг. → 6 СО2 + 44 Н2О + 38 АТФ
След като се синтезира пируват, не е задължително да се превърне в ацетил-SКоА. Съществено значение има способността на пирувата да се карбоксилира до оксалоацетат, тази реакция е активна в черния дроб. Наличието на излишен оксалоацетат стартира реакциите от цикъла на трикарбоновите киселини (фиг.157).
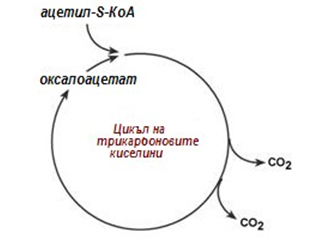
Фигура 157. Роля на оксалоацетата за аеробния метаболизъм
При анаеробно окисление на глюкозата, пирувата се превръща в лактат. Лактата е краен продукт, който не се метаболизира, единствено може да се окисли отново до пируват. В микробиологията анаеробната гликолиза е известна като млечна ферментация. Общото уравнение на анаеробната гликолиза, може да се представи като:
С6Н12О6 + 2 АДФ + 2 Фнеорг. → 2 Лактат + 2 Н2О + 2 АТФ
Много клетки в организма имат способност анаеробно да окисляват глюкозата. Например за еритроцитите, това се явява единствен източник на енергия. Клетките на скелетната мускулатура при мощна, интензивна и бърза работа (бягане на къса дистанция – спринт) използват безкислородното оксиление на глюкозата.
11.2. Гликолиза
11.2.1. Обща характеристика и локализация
В клетките на прокариотните гликолизата протича в цитоплазмата. В еукариотните организми десетте фермента, катализиращи разпада на глюкозата до пируват се намират в цитозола (фиг.158).
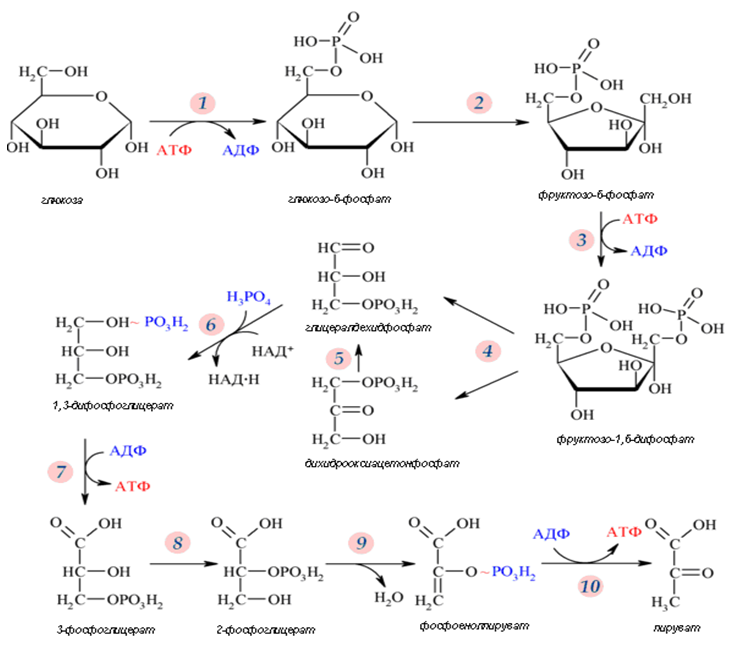
Фигура 158. Обща схема на гликолизата
Гликолитичният път представлява серия от 10 последователни реакции, всяка от които се катализира от отделни ензими. Процеса условно се разделя на два етапа:
Ø Първи етап (подготвителен) – включва първите три реакции, които протичат на ниво хексозни молекули. Реактивни са само цикличните форми. Хексозната молекула на глюкозата се подготвя за разграждане, като се активира за сметка на разграждането на 2 молекули АТФ, завършва с разпадане до 2 молекули глицералдехид-3-фосфат.;
Ø Втори етап (окислителен) - протича НАД-зависимо окисление, съпроводено със синтез на АТФ.
11.2.2. Химизъм на гликолизата
В подготвителната фаза на гликолизата глюкозата се фосфорилира от хексокиназа (глюкокиназа в черния дроб) до глюкозо-6-фосфат. Последният се изомеризира до фруктозо-6-фосфат от хексозофосфат изомераза. Фруктозо-6-фосфатът се фосфорилира от фосфофруктокиназа до фруктозо-1,6-дифосфат. Тези реакции консумират 2 молекули АТФ за 1 молекула глюкоза. С получаването на фруктозо-1,6-дифосфат завършва подготвителната фаза в гликолизата.
Първа реакция. Активирането на глюкоза до глюкозо-6-фосфат с участието на АТФ е фосфотрансферазна реакция (реакция 1). За протичане 2+ на реакцията е необходимо наличие в средата на Mg йони, с които молекулата АТФ се свързва комплексно. Пренася се фосфатен радикал от макроергично на нормоергично ниво. Част от енергията, отделена при разграждане макроергичната връзка на АТФ се включва в новосъздадената естерна връзка, а остатъкът се разсейва като топлина. Ензимът, катализиращ фосфорилирането на глюкоза, се нарича хексокиназа. Известни са 4 изоензими. Тук се изразходва 1 макроергична връзка. Реакцията е необратима и е първа ключова реакция на гликолизата.
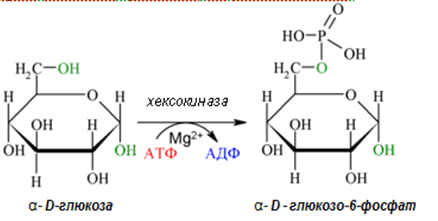
Фигура 159. Активиране на глюкоза до глюкозо-6-фосфат с участието на АТФ (реакция 1 - първа ключова реакция на гликолизата).
Фосфорилирането на глюкозата има две цели:
Ø Първо - плазматичната мембрана, която е проницаема за неутралната молекула на глюкозата не пропуска отрицателно заредените молекули на Г-6-Ф, фосфорилираната глюкоза се оказва затворена вътре в клетката.
Ø Второ - при фосфорилирането глюкозата преминава в активна форма, способна да участва в биохимически реакции и да се включва в метаболитни цикли.
Втора реакция. Алдозата глюкозо-6-фосфат се превръща в кетозата фруктозо-6-фосфат от глюкозо-фосфат изомераза (реакция 2). Тази изомеразна реакция е напълно обратима и протича като вътрешно - молекулна оксидо-редуция. На този етап в гликолизата може да се включва чрез фосфорилиране и фруктоза.
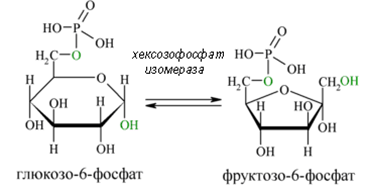
Фигура 160. Превръщане на глюкозо-6-фосфат в кетозата фруктозо-6-фосфат от ензима глюкозо-фосфат изомераза (реакция 2).
Трета реакция. След това фруктозо-6-фосфат се активира допълнително до фруктозо-1,6-дифосфат от фосфофруктокиназа за сметка на разграждане на още една молекула АТФ (реакция 3). Фосфофруктокиназата е фосфотрансфераза като хексокиназата. Фосфофруктокиназата е най- важният регулаторен алостеричен ензим за гликолиза. Докато хексокиназата участва и в други обменни пътища, фосфофруктокиназата е първият специфичен само за гликолизата ензим.
С получаването на фруктозо-1,6-дифосфат завършва подготвителната фаза в гликолизата. Консумират се 2 молекули АТФ за 1 молекула глюкоза.
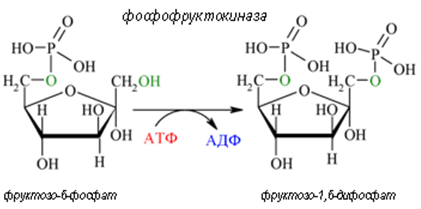
Фигура 161. Фруктозо-6-фосфат се активира допълнително до фруктозо-1,6-дифосфат от ензима фосфофруктокиназа (реакция 3).
Четвърта реакция. Следва обратимо алдолно разцепване на образуваният фруктозо-1,6-дифосфат на две триози (реакция 4). Фруктозо- 1,6-дифосфата се разгражда под действие на ензима алдолаза до триозите глицералдехид-3-фосфат и дихидроксиацетон фосфат, които се превръщат едно в друго под действие на триозофосфат изомераза (реакция 5).
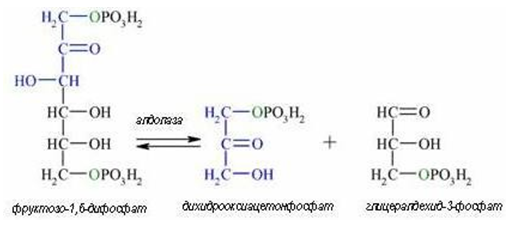
Фигура 162. Фруктозо-1,6-дифосфата се разгражда от алдолаза до триозите глицералдехид-3-фосфат и дихидроксиацетон фосфат (реакция 4).
Пета реакция. В резултат на четвъртата реакция се образува дихидроксиацетонфосфат и глицералдехид-3-фосфат, при това под действието на фосфотриозоизомеразата, дихидроксиацетонфосфата почти веднага преминава в глицералдехид-3-фосфат (реакция 5).
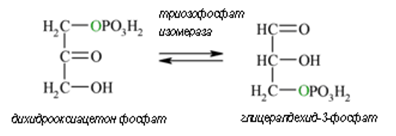
Фигура 163. Глицералдехид-3-фосфат и дихидроксиацетон фосфат се превръщат едно в друго под действие на триозофосфат изомераза (реакция 5).
Шеста реакция. Глицералдехид-3-фосфат се изтегля в следващата реакция, което улеснява превръщането на ДХАФ в глицералдехид-3-фосфат. В реакция 6, алдехидната група на глицералдехид-3-фосфат се окислява до карбоксилна. Отделената енергия се акумулира в ацил-фосфатна макроергична връзка. Получава се макроергичен продукт 1,3- дифосфоглицерат. Ензимът глицералдехид-3-фосфат дехидрогеназа е анаеробна дехидрогеназа, действаща съвместно с НАД+, който се редуцира.
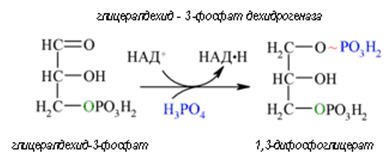
Фигура 164. Глицералдехид-3-фосфат се окислява до 1,3-дифосфоглицерат (реакция 6).
Седма реакция. 1,3-дифосфоглицерата, съдържа макроергична връзка, ензима фосфоглицераткиназа пренася остатъка на фосфорната киселина на молекула АДФ (реакция 7) — образува се молекула АТФ. Това е първата реакция на субстратно фосфорилиране. Синтезират се 2 молекули АТФ (по една на всяка молекула 1,3-дифосфоглицерат), за протичането на реакцията се изисква присъствие в цитозола на АДФ, при излишък в клетката на АТФ (и недостатък на АДФ) скоростта се намалява. Тъй като АТФ, не се подлага на метаболизъм, не се депонира в клетката а просто се разрушава, то тази реакция е важен регулатор на гликолизата.
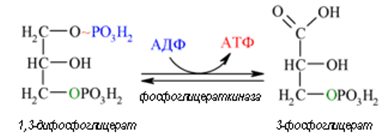
Фигура 165. Субстратно фосфорилиране (реакция 7).
Осма реакция. В реакция 7, глицерат киназа пренася фосфатната група на високо макроергично ниво (от 1,3-дифосфоглицерат върху АДФ). Натрупаната енергия в ацил-фосфатната връзка се пренася върху АДФ и се синтезира АТФ (субстратно фосфорилиране), а 1,3-дифосфоглицерат се превръща в 3-фосфоглицерат. Последният е субстрат за ензима фосфоглицерат мутаза, която го превръща обратимо в 2-фосфоглицерат (реакция 8).
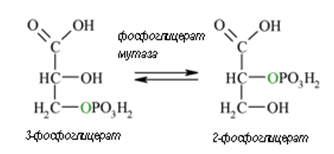
Фигура 166. Фосфоглицерат мутаза превръща обратимо 3-фосфоглицерат в 2- фосфоглицерат (реакция 8).
Девета реакция. Реакция 9, т.нар. енолазна реакция, е също пример за субстратно фосфорилиране. Тук 2-фосфоглицерат се превръща без участие на външна редокс-система в макроергичния продукт фосфоенолпируват (ФЕП). Отделянето на вода протича като вътрешномолекулна оксидоредукция, съпроводена с отделяне на енергия. Тази енергия се акумулира в енолфосфатната връзка на ФЕП, предшественик на АТФ.
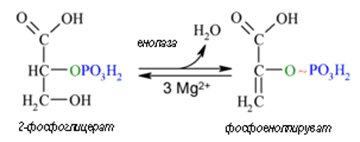
Фигура 167. 2-фосфоглицерат се превръща без участие на външна редокс-система в макроергичния продукт фосфоенолпируват (ФЕП) (реакция 9).
Десета реакция. В реакция 10 пируват киназа превръща ФЕП в пируват (фиг.168). Реакцията е необратима и силно екзергонична (∆Go' = - 61.9 kJ/mol).
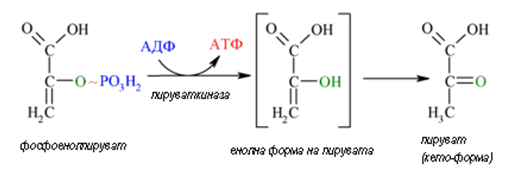
Фигура 168. Пируват киназа превръща ФЕП в пируват (реакция 10).
Част от акумулираната енергия във ФЕП се ползва за синтеза на АТФ – пренася се фосфатен радикал на макроергично ниво от ФЕП върху АДФ, а останалата част се разсейва като топлина. За високата стойност на ∆Go' допринася главно тавтомеризирането на енол-пируват в неговата кето- форма.
В глицераткиназната и пируваткиназната реакция се получават 4 молекули АТФ за 1 молекула глюкоза и общата енергетична равносметка при анаеробни условия е 2 молекули АТФ за молекула глюкоза.
В отсъствие на кислород (анаеробна гликолиза), пируват се редуцира до лактат, съпроводено с окисление на НАДН до НАД+. Така се регенерира НАД+, необходим за окислителното фосфорилиране на глицералдехид-3-фосфат. Лактат се получава както в усилено работещи мускули, така и при млечнокисела ферментация. При алкохолна ферментация пируват през ацеталдехид дава етанол.
Реакциите, катализирани от хексокиназа, фосфофруктокиназа и пируваткиназа са необратими. Тези три ензима са регулаторните ензими на гликолизата. Главният регулаторен ензим фосфофруктокиназа се инхибира алостерично от АТФ и цитрат, а АМФ, АДФ и фруктозо-2,6-дифосфат действат като активатори.
Гликолизата е свързана с цитратния цикъл посредством окислителното декарбоксилиране на пируват до ацетил-КоА, лигазното карбоксилиране на пируват до оксалацетат и редуктивното карбоксилиране на пируват до малат. Последните две реакции са анаплеротични (попълват резервоара от оксалоацетат и малат).
11.3. Биосинтеза на въглехидратите при автотрофните организми
Всички организми мога да бъдат разделени на две големи групи:
Ø автотрофни;
Ø хетеротрофни.
Автотрофните притежават способността да синтезират органични вещества от неорганични вещества.
Хетеротрофните изграждат своите органични вещества от готови органични съединения, които само преработват. Хетеротрофните организми живеят за сметка на автотрофните. За да се осъществи синтеза на органични вещества е необходима енергия. В зависимост от използваните съединения и източниците на енергия се различават следните основни типове въглеродно хранене (табл.6).
Таблица 6. Видове въглеродно хранене
Вид въглеродно хранене |
Източник на въглерод |
Източник на водород |
Източник на енергия |
Хетеротрофно хранене |
органични съединения |
органични съединения |
органични съединения |
Автотрофно хранене (3 вида) 1) фотосинтеза |
СО2 |
Н2О |
светлинни кванти |
2) фоторедукция (бактериална фотосинтеза) |
СО2 |
H2S, H2 |
светлинни кванти |
3) хемосинтеза (от хемосинтезиращи бактерии) |
СО2 |
H2S, H2, Н2О, NH3 |
химична енергия, която се получава при разграждане на различни съединения и от различни химични реакции. |
При автотрофните организми, синтезата на глюкоза се извършва чрез процесите:
Ø Фотосинтеза;
Ø Хемосинтеза.
11.3.1. Същност на фотосинтезата
Основен източник на енергия за почти всички живи организми на з емята е слънцето. Слънчевата енергия се въвежда в биосферата чрез процес известен като фотосинтеза, който се извършва от растенията, алгите и някои видове бактерии. Ежегодно чрез фотосинтеза се фиксират около 1011 t въглерод в органични съединения, което означава съхранение на 1018 kJ от слънчевата енергия. Това количеството енергия многократно превишава добиваното от другите източници (въглища, нефт, природен газ).
Фотосинтезата е биологичен процес, характерен за зелените растения и някои микроорганизми (цианобактерии), може да се дефинира като физико- химичен процес, при който фотосинтезиращите организми използват светлинната енергия за да извършват синтез на органични вещества.
Фотосинтезата е единен процес, но от биохимична гледна точка може да бъде разделен на две фази:
Ø Светлинна;
Ø Тъмнинна.
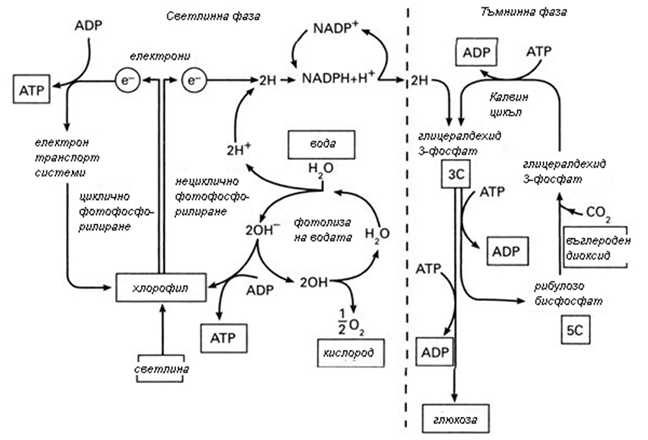
Фигура 169. Светлинна и тъмнинна фаза на фотосинтезата
Светлинна фаза. Протича в хлоропластите, само в присъствие на светлина, с прякото участие на хлорофил и други спомагателни пигменти, свежда се до получаване АТФ и НАДРН, като едновременно с това се отделя кислород. Осъществява се чрез поглъщане на слънчева енергия (кванти) от хлорофила и другите пигменти на фотосинтезиращите клетки.
Слънчевата енергия е форма на електромагнитна енергия, която пътува под формата на електромагнитни вълни. Разстоянието между две съседни вълни се нарича дължина на вълната. Диапазона от различните видове електромагнитна енергия, от късите гамма лъчи до дългите радио вълни се обозначава като електромагнитен спектър (фиг.170).
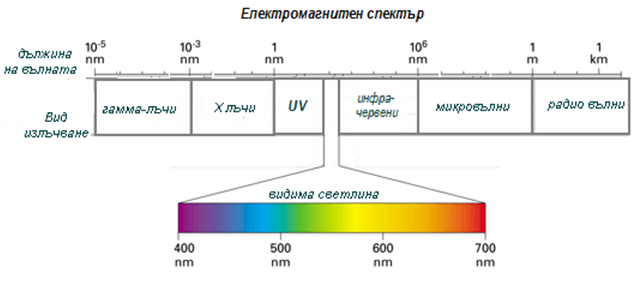
Фигура 170. Електромагнитен спектър (вид излъчване и дължина на вълната)
Различните видове излъчвания имат различна дължина на вълната, като с по-късите дължини на вълната притежават повече енергия. Видимата светлина, са електромагнитни вълни с различна дължина на вълната от 400 nm до 700nm, които се виждат с различен цвят и са само малка фракция от електромагнитния спектър.
Пигментите в хлоропластите са субстанции оцветени с различен цвят, поради съдържание на специфични химични съединения. Ролята на тези пигменти, особено на хлорофила е да поглъща светлината, което протича особено интензивно в две области: 400÷500 nm и 600÷700 nm.
Светлопоглъщащите пигменти са събрани в ансамбли, наречени фотосистеми, изградени от 200 молекули хлорофил и 50 молекули каротеноиди. Светлината попадаща върху пигментите може да се абсорбира, премине или отрази (фиг.171).
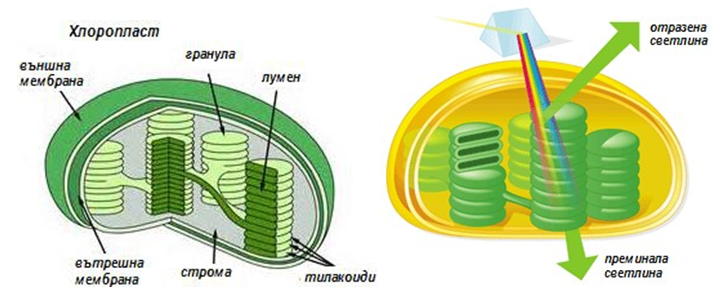
Фигура 171. Структура на хлоропластите и поведение на светлината попаднала върху тях
Пигментите на хлоропластите не абсорбират светлината със зелен цвят от видимия електромагнитен спектър, повечето светлина със зелен цвят преминава през листа или се отразява, поради което листата на растенията имат зелен цвят (фиг.171).
В хлоропластите на растителните клетки и по-точно в тилакоидните мембрани, хлорофила и другите пигментни молекули са подредени в кластери наречени фотосистеми (I и II), и съдържат стотици пигментни молекули. Фотосистемите са събиращи светлината единици в тилакоидните мембрани.
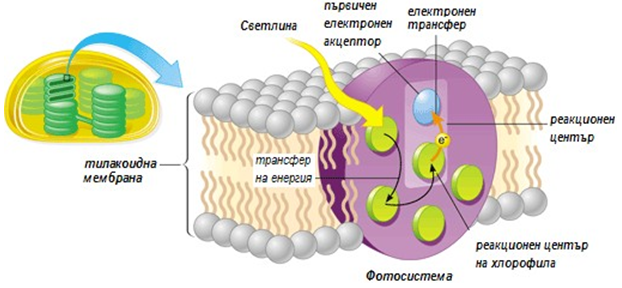
Фигура 172. Реакционен център на хлорофила разположени в тилакоидните мембрани на хлоропластите
Всеки път, когато пигментната молекула абсорбира слънчева енергия (квант), един от електроните на пигментната молекула поглъща енергията и преминава в нестабилно състояние. Почети веднага, възбуденият електрон се връща обратно в основното си състояние като прехвърля енергията на съседна молекула, енергията прескача от молекула на молекула докато не достигне до така наречения реакционен център на фотосистемата (фиг.172).
Всяка фотосистема притежава стотици пигментни молекули подредени в комплекси наречени антена (фиг.173). Системата на антената е изградена от стотици пигментни молекули (основно хлорофил или бактериохлорофил, и каротеноиди), които са прикрепени към протеини във фотосинтетичните мембрани и обслужват специален протеинов комплекс известен като реакционен център. Всеки път, когато фотон удари пигментна молекула, енергията се прехвърля от молекула на молекула, докато не достигне реакционния център.
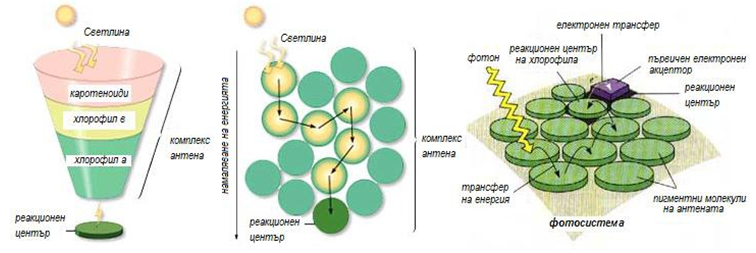
Фигура 173. Реакционен център на фотосистема
Познати са два вида хлорофил (а и в), съдържат се в растенията и два вида бактериохлорофил (а и в), които са компоненти на фотосинтезиращите микроорганизми. Те се различават единствено по състав на R1, R2, R3 и R4. Към спомагателните компоненти се отнасят каротиноиди, оцветени в жълто или червено.
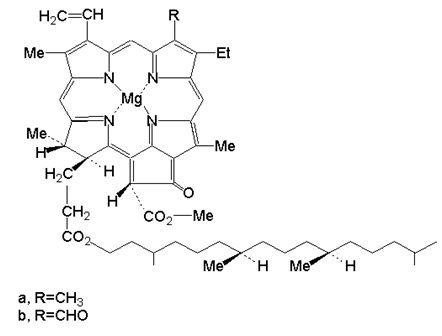
Фигура 174. Структура на хлорофил а и в
При растенията, две фотосистеми са включени в светлинните реакции, които функционират взаимосвързано (фиг.175):
Ø Фотосистема I - изградена предимно от хлорофил а, който може да бъде възбуден от светлинни лъчи с дължина на вълната – 700 nm;
Ø Фотосистема II - преобладава хлорофил в и поглъща максимално светлина с дължина на вълната 680 nm.
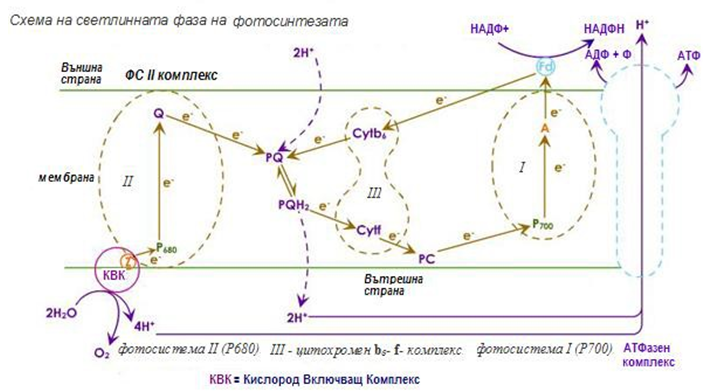
Фигура 175. Схема на светлинната фаза на фотосинтезата
Светлинните реакции протичат като конвертират светлинната енергия в енергия на химичните връзки (фиг.176).
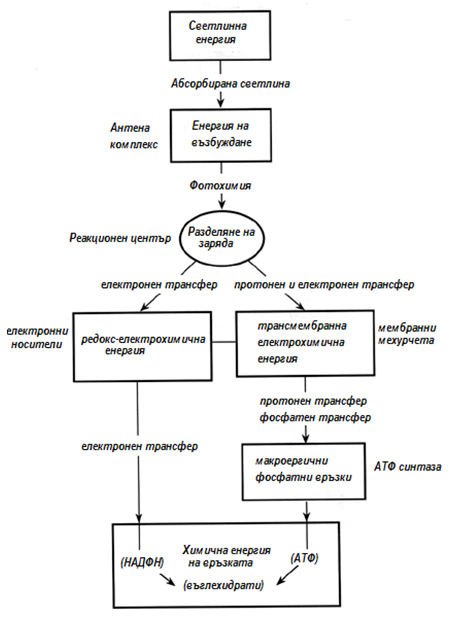
Фигура 176. Трансформиране на енергията при фотосинтезата
Фотосистема I (Р700). При попадане на фотон (hv) върху Р700 , нейният реакционен център преминава във възбудено състояние Р700*. Отделя се електрон, който се пренася през верига (I) от междинни носители – А0 (мономер на хлорофил а, А1-филохинон и Fx, Fa, Fb – три железно-серни белтъка) до разтворимия фередоксин (Фд). Фередоксин (Фд) се редуцира и участва в процеса на получаване на НАДФH от НАДФ+, с участието на фередоксин-НАДФ-редуктазата.
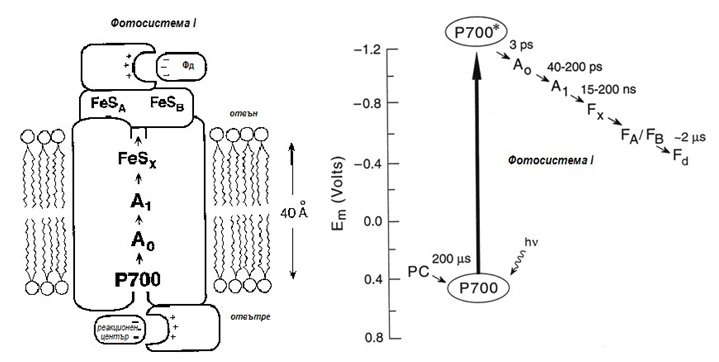
Фигура 177. Фотосистема I (Р700). При попадане на фотон (hv) върху Р700 , нейният реакционен център преминава във възбудено състояние Р700*. Отделя се електрон, който се пренася през верига (I) от междинни носители – А0 (мономер на хлорофил а, А1-филохинон и Fx, Fa, Fb – три железно-серни белтъка) до разтворимия фередоксин (Фд).
Циклично фосфорилиране. Освен тази електрон-пренасяща верига съществува още една, по която възбудения и напуснал Р700* елект рон се връща обратно в Р700. Тя включва междинните системи – А0, А1, Fx, Fa, цитохром b6, цитохром f и пластоцианин. При движението си електронът понижава постепенно своята енергия и преминава през участък (цитохром b6 и цитохром f), в който молекула АДФ се фосфорилира до АТФ за сметка на част от неговата енергия. Този процес е известен като циклично фосфорилиране, при което се получава само АТФ, но не и НАДФН.
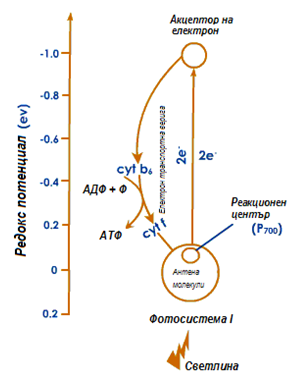
Фигура 178. Схема циклично фосфорилиране
Механизмът, чрез който се отклоняват електроните от фотосистема I в една или друга посока, не е достатъчно добре проучен.
Фотосистема II (P680). При отделянето на електрон от Р700 се образува “електронна дупка”, която се попълва от Р680. Освен това за пълната редукция на НАДФ+ освен електрони са необходими и протони, които се набавят от фотолизата на водата, осъществявана от Р680.
Нециклично фосфорилиране. При попадане на светлинни кванти върху фотосистема II, лъчи с дължина на вълната 680 nm, се поглъщат и възбуждат системата Р680*. Отделя се електрон, който преминава през верига от електронни носители – феофитин а (Pheo a), пластохинон А (РQA), пластохинон B (РQB), цитохром b6, цитохром f и пластоцианин, който е непосредствен донор на електрони за електронната дупка във фотосистема
I. При придвижването си в тази верига, електроните отдават част от
енергията си в участъка между цитохром b6 и цитохром f , и се синтезира АТФ от АДФ. Този процес е известен като нециклично фосфорилиране, тъй като електронът се отделя от фотосистема II и постъпва във фотосистема I.
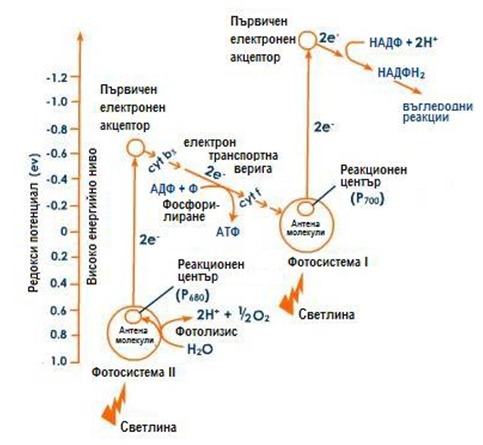
Фигура 179. Схема на нециклично фосфорилиране
Тъмнинна фаза на фотосинтезата (цикъл на Калвин). Метаболитния път, чрез който растенията синтезират въглехидрати от СО2, се означава като тъмнинна фаза, тъй като може да протече без светлина.
Условно е разделен на два етапа:
Ø Продуктивен. През първият етап от 6 молекули СО2 се синтезира молекула глюкоза с участието на 6 молекули рибулозо-1,5-дифосфат, 6 молекули вода, 12 молекули НАДФН и 12 молекули АТФ.
Ø Възстановителен. През втория етап започва серия от реакции, подобни на тези от пентозофосфатния път, при което се възстановяват 6 молекули рибулозо-1,5-дифосфат, за което се изразходват нови 6 молекули АТФ.
Редукция на СО2. Процесът започва с присъединяването на СО2 към рибулозо-1,5-дифосфата. Реакцията се катализира от рибулозо-дифосфат карбоксилаза. Получава се шествъглеродно неустойчиво съединение, което се разгражда бързо на две молекули фосфоглицерат с участието на вода. Това е най-типичния процес на фотосинтезата и е необратим.
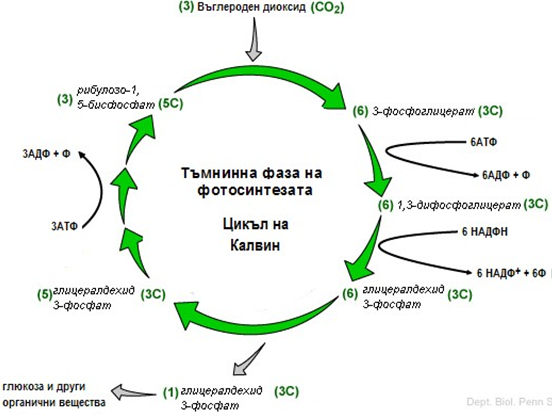
Фигура 180. Схема на Цикъла на Калвин
Реакции от гликолитичния път във фотосинтезата. Следващите реакции са стъпала на гликолитичната верига, но протичат в обратна посока, при фотосинтезата. За целта се използва енергията, уловена в светлинната фаза под формата на АТФ и НАДФН.
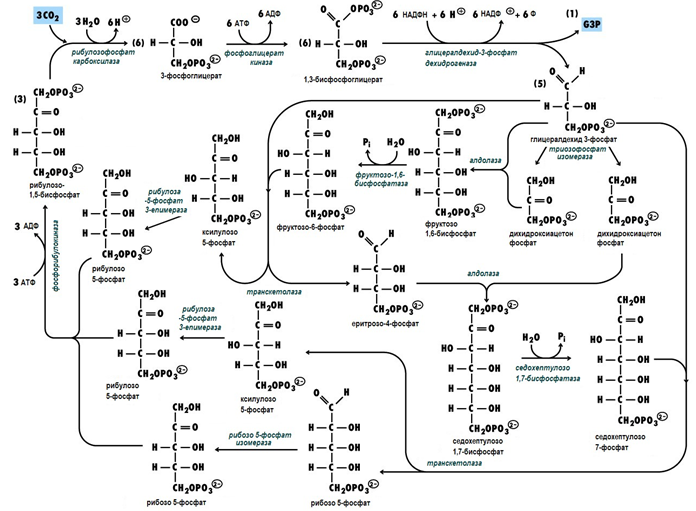
Фигура 181. Химизъм на реакциите от Цикъла на Калвин
Реакции от пентозофосфатния цикъл във фотосинтезата. Реакциите познати от пентозофосфатния цикъл и се катализират от транскетолаза, алдолаза, рибозо-5-фосфат изомераза и рибулозо-5-фосфат епимераза.
Три от ензимите са типични само за фотосинтезата рибулозодифосфат карбоксилаза, седохептулозодифосфатаза, фосфорибулокиназа и нямат еквивалент в животинските тъкани.
Сумарно уравнение на цикъла на Калвин. Сумарното уравнение на цикъла на Калвин има следния вид:
6СО2 + 12Н2О + 12НАДФН + 18АТФ → С6Н12О6 + 12НАДФ+ + 18АДФ + 18Н3РО4
Дълго време е считано, че при фотосинтезата се образуват само въглехидрати, които след това метаболизират в други съединения чрез реакции, несвързани с фотосинтезата. Експериментално е доказано, че от междинни продукти могат да се получат мастни киселини и АК. Например серин се получава чрез пряко аминиране на 3-фосфоглицерат, който лесно може да премине във фосфоенолпируват и след това в пируват. Той може да встъпи в разнообразни реакции, водещи до получаване на мастни киселини, органични киселини, аланин и др.
11.3.2. Хемосинтеза
Органични вещества се синтезират не само по пътя на фотосинтезата в зелените растения, но и от бактерии, които не съдържат хлорофил. Те усвояват СО2, като използват енергията, образуваща се при окисление на различни неорганични съединения: Н2S, S, H2, NH3, HNO2, оксиди на желязото и мангана. Този процес на синтеза на органични съединения, протичащ за сметка на енергията, отделена при оксиление на неорганични съединения, носи наименованието хемосинтеза.
При хетеротрофните организми, синтеза на глюкоза от невъглехидратни предшественици е процес наречен – глюконеогенеза.
11.4. Глюконеогенеза
В организма винаги е необходимо да се синтезира глюкоза:
Ø за еритроцитите глюкозата е единствен източник на енергия;
Ø нервната тъкан употребява около 120 гр. глюкоза в денонощие и тази стойност не зависи от интензивността на работа. Единствено в
екстремни ситуации (гладуване), тя е способна да получава енергия от невъглеродни източници.;
Ø глюкозата изпълнява водеща роля в поддържане необходимата концентрация на метаболитите от цикъла на трикарбоновите киселини (оксалоацетат).
В определени ситуации, при диета с ниско съдържание на въглехидрати, гладуване, дълга продължителна физическа работа, т.е когато глюкозата в кръвта е изразходена и настъпва хипогликемия, организма е длъжен да има възможност да синтезира глюкоза и да нормализира нейната концентрация в кръвта. Това се осъществява чрез глюконеогенезата.
По определение глюконеогенез е синтез на глюкоза от невъглехидратни компоненти: лактат, пируват, глицерол, кетокиселини от цикъла на Кребс и други кетокиселини получени от трансаминиране на аминокиселини.
Необходимостта от глюконеогенезата в организма се демострира от два цикъла – глюкозо-лактатен и глюкозо-аланинов.
11.4.1. Глюкозо-лактатен цикъл (цикъл на Кори)
Глюкозо-лактатния цикъл е цикличен процес, обединяващ реакциите от гликонеогенезата и реакциите на анаеробната гликолиза. Глюконеогенезата протича в черния дроб, субстрат за синтезата му е лактат, постъпващ в организма от еритроцитите или мускулната тъкан.
В еритроцитите млечна киселина се образува непрекъснато, тъй като анаеробната гликолиза се явява единствен способ за добиване на енергия.
В скелетните мускули натрупване на лактат настъпва от протичане на анаеробна гликолиза, при интензивна и субмаксимална мощност на работа, при което вътреклетъчното рН се понижава до 6,3 – 6,5. Премахването на млечната киселина може да се осъществи единствено като се превърне в пируват. Мускулната клетка не е способна да осъществи тази реакция, поради специфичността на ензима ЛДГ-5, който превръща пирувата в лактат, но не катализира обратната реакция. Лактата преминава през мембраната на клетката навън по концентрационен градиент. Голяма част от лактата в кръвта се улавя от хепатоцитите, окислява се до пируват и постъпва в глюконеогенезата. Глюкозата образувана в черния дроб от хепатоцитите или се използва от тях, или се връща в мускулните клетки като възстановява запасите от гликоген, или се използва от клетките на другите органи.
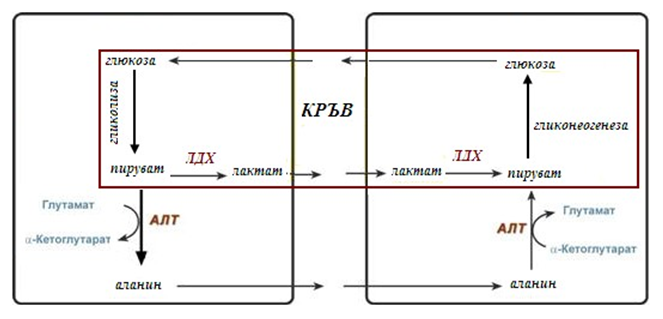
Фигура 182. Глюкозо-лактатен и глюкозо-аланинов цикъл
11.4.2. Глюкозо-аланинов
Пирувата е токсичен за клетката и клетката се стреми да се освободи от него. Тъй като клетъчната мембрана е непроницаема за пирувата, то обезвреждането на пируват, може да се осъществи чрез превръщането му:
Ø в лактат;
Ø в ацетил-SКоА;
Ø в аланин;
Ø в оксалоацетат.
Цел на глюкозо-аланиновия цикъл е премахването на пирувата и също така на излишния азот от мускулите.
При мускулна работа и в покой, в мускулните клетки се разграждат белтъци и образуваните аминокиселини се трансаминират с α-кетоглутарат. Полученият глутамат взаимодейства с пирувата. Образува се аланин, който е транспортна форма на азота и пирувата от мускулите до черния дроб. В хепатоцитите протича обратната реакция на трансаминиране, аминогрупата се предава на орнитиновия цикъл за синтез на уреа, пирувата се използва за синтез на глюкоза.
Освен при мускулна работа, глюкозо-аланиновия цикъл се активира при гладуване, когато мускулните белтъци се разпадат и много аминокиселини се използват в качеството на източник на енергия, а техния азот трябва да се транспортира до черния дроб.
11.4.3. Химизъм на глюконеогенезата
Глюконеогенезата е синтез на глюкоза от невъглехидратни компоненти: лактат, пируват, глицерол, кетокиселини от цикъла на Кребс и други кетокиселини получавани от аминокиселини. Всички аминокиселини, с изключение на кетогенните левцин и лизин, участват в синтезата на глюкоза. Освен получаването на глюкоза, глюконеогенезата обезпечава премахването на шлаките – лактата, който постоянно се образува от еритроцитите или от мускулните клетки, и на глицерола, продукт от липолизата в мастната тъкан.
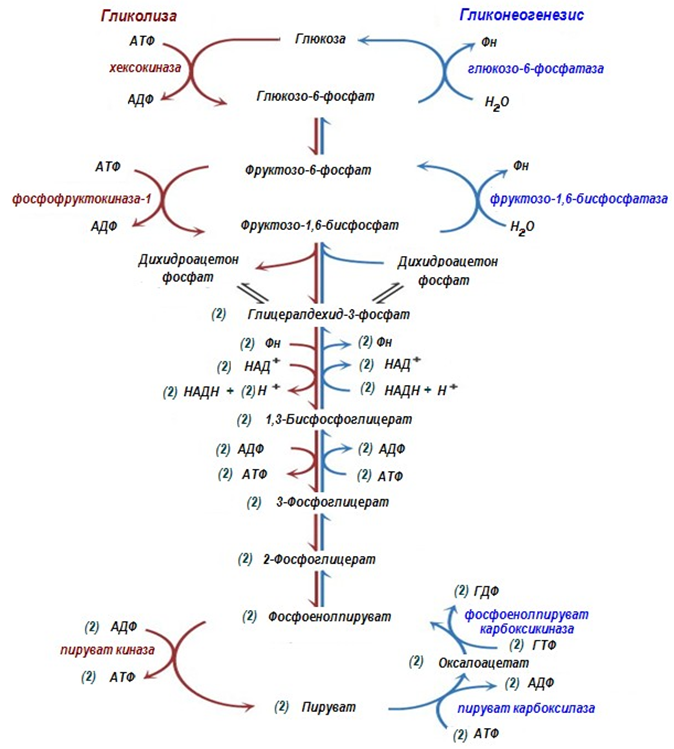
Фигура 183. Гликолиза и глюконеогенеза
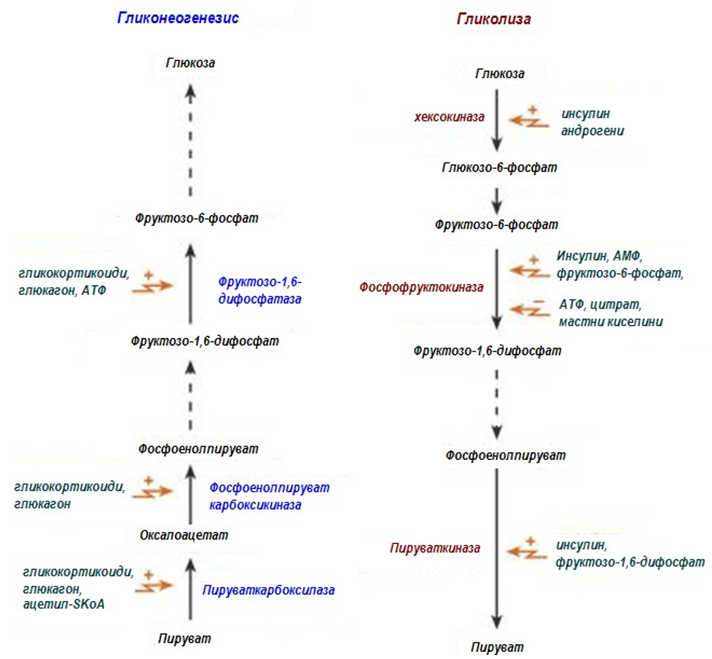
Глюконеогенезата е път, обратен на гликолизата. Всички обратими реакции от гликолизата са част от глюконеогенеза в обратна посока на тази в гликолизата. Само трите необратими реакции в гликолизата, при които ∆Go' има високи отрицателни стойности, се заместват от реакции, които правят синтезата на глюкоза термодинамично възможна.
В гликолизата има три необратими реакции:
Ø пируваткиназната (десета),
Ø фосфофруктокиназната (трета) и
Ø хексокиназната (първа).
В тези реакции протича освобождаване на енергия за синтез на АТФ. Ето защо в обратния процес възниква енергетическа бариера, която клетката заобикаля с помощта на допълнителни реакции.
Глюконеогенезата включва всичките обратими реакции на гликолизата, и обходни пътища за необратимите, т.е. той не повтаря напълно реакциите на окисление на глюкозата. Неговите реакции са способни да протекат във всички тъкани, освен последната глюкозо-6-фосфатазната реакция, която протича единствено в черния дроб и бъбреците.
Глюконеогенезата действа в черния дроб и в по-малка степен в бъбреците. Включва се, когато храната не съдържа глюкоза и са изчерпани запасите от гликоген в черния дроб. Ето защо, може да се каже, че глюконеогенез протича единствено в тези два органа.
Обход на десета реакция на гликолизата (фиг. 183, 184, 185). На този етап на глюконеогенезата работят два ключови ензима – в митохондриите пируваткарбоксилаза и в цитозола фосфоенолпируват-карбоксикиназа. В химически план обходния път на десетата реакция изглежда достатъчно просто:
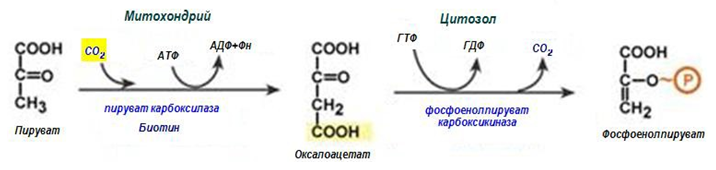
Фигура 184. Обход на десета реакция на гликолизата
Пируваткарбоксилазата се намира в митохондрия, а фосфоенолпируват- карбоксикиназата – в цитозола. Допълнителен проблем е непроницаемостта на митохондриалната мембрана за оксалоацетат. През мембраната може да премине малат, предшественик на оксалоацетат в ЦТК. Реално процеса протича по-сложно:
1. В цитозола пируват може да се образува при окисление на млечна киселина и от реакциите на трансаминиране на аланина. След това пируват със симпортен пренос с йоните Н+, движещи се по протонния градиент, прониква в митохондриите. В митохондриите пируваткарбоксилазата превръща пируват в оксалоацетат. Тази реакция протича в клетката постоянно, явява се анаплеротическа (допълваща) реакция на ЦТК.
2. Оксалоацетат може да се превърне във фос фоенолпируват, но за това трябва да попадне в цитозола. Ето защо протича реакция на възстановяване оксалоацетат в малат при участие на малатдехидрогеназа. В резултат се натрупва малат, който излиза в цитозола и тук се превръща обратно в оксалоацетат. Малатдехидрогеназната реакция от ЦТК протича поради излишък на НАДН в митохондриите. НАДН постъпва от
β‑окислението на мастните киселини, който се активира при условия на недостатъчно глюкоза в хепатоцитите.
3. В цитоплазмата фосфоенолпируват-карбоксикиназа осъществява превръщане на оксалоацетат в фосфоенолпируват, за реакцията е необходима енергия ГТФ. От молекулата се отделя този въглерод, който се е присъединил.
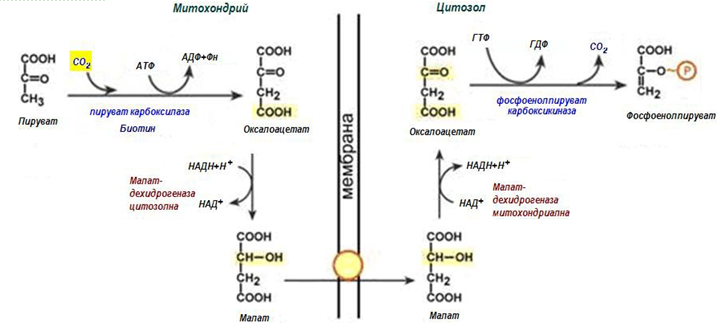
Фигура 185. Схема на обходния път на десета реакция на гликолизата
Обход на трета реакция на гликолизата (фиг.183, 186). Второ препятствие по пътя на синтезата на глюкоза е фосфофруктокиназната реакция, която се преодолява с помощта на ензима фруктозо-1,6- дифосфатаза. Този ензим се съдържа в бъбреците, черния дроб, напречно- набраздената мускулатура. Тези тъкани са способни да синтезират фруктозо-6-фосфат и глюкозо-6-фосфат.
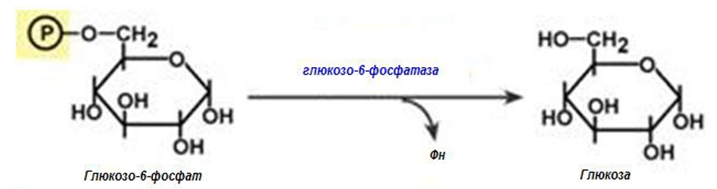
Фигура 186. Обход на трета реакция на гликолизата
Обход на първа реакция на гликолизата. Последната реакция от глюконеогенезата се катализира от глюкозо-6-фосфатаза. Този ензим присъства единствено в черния дроб и бъбреците, следователно само в тези тъкани може да се синтезира глюкоза.
Фигура 187. Обход на първа реакция на гликолизата
11.4.4. Регулиране на глюконеогенезата
Противоположните пътища гликолиза и глюконеогенеза не протичат едновременно in vivo, а се регулират реципрочно в зависимост от нуждите на организма.
Регулаторните ензими за глюконеогенеза са:
Ø пируват карбоксилаза и ФЕП карбоксикиназа;
Ø фруктозо-1,6-бисфосфатаза;
Ø глюкозо-6-фосфатаза.
Катализираните от тях реакции са екзергонични и необратими.
Фигура 188. Регулация на гликолизата и глюконеогенезата
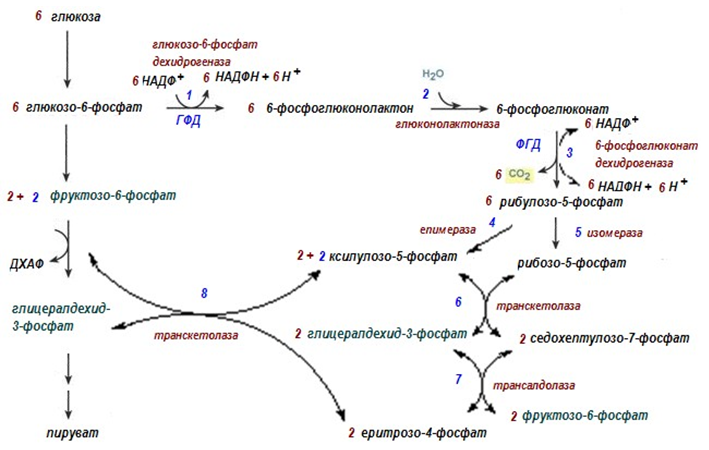
11.5. Пентозофосфатен път (ПФП)
11.5.1. Обща характеристика на ПФП
В клетките непрекъснато се синтезира белтък, за което са необходими рибонуклеинови киселини. За синтезата на нуклеиновите киселини и по- точно на пурини и пиримидини е необходим рибозо-5-фосфат. При клетки, които се подготвят за делене протича интензивна синтеза на ДНК, за което са необходими дезоксорибонуклеотиди, които се образуват с участието на НАДФН. Молекулите на НАДФН се използват при синтез на:
Ø мастни киселини;
Ø холестерол;
Ø глутаминова киселина от α-кетоглутарат;
Ø при системата на антиоксидантна защита на клетките от окислението на свободните радикали (еритроцити).
Процеса, който обезпечава клетките едновременно с рибози и НАДФН е пентозофосфатния път (ПФП).
ПФП носи названието директно (пряко) окисление на глюкозата, тъй като още първите две реакции са окислително-редукционни процеси. Пентозофосфатният път е известен и като фосфоглюконатен път, първата реакция е окисление на глюкозо-6-фосфата до глюконат. Окислението започва на ниво хексози, а не на ниво триози.
Най-активно реакциите на ПФП протичат в цитозола на клетките на черния дроб, мастната тъкан, еритроцитите, млечните жлези при лактация и много по-слабо в скелетната мускулатура.
Фигура 189. Обща схема на реакциите на ПФП
ПФП окислява глюкозата и при определени условия може напълно да я разгради до СО2 и Н2О, но се класифицира като анаболитен път, който използва 6-те въглеродни атома на глюкозата, за да образува въглехидрати с 5 въглеродни атоми.
Основни функции на ПФП са:
Ø образува редуциран НАДФH, необходим за биосинтези в клетката;
Ø снабдява клетката с рибозо-5-фосфат за синтеза на нуклеинови киселини;
Ø чрез специфични трансферазни реакции може да образува субстрати за гликолизата и гликонеогенезата;
Ø при пълно разграждане на 1 молекула глюкоза се получават 12 мола НАДФН, ако пътят се свърже с окислителното фосфорилиране в дихателната верига би се образувал 36 мол АТФ, както при пълното разграждане на глюкозата в гликолизата и ЦТК. Този метаболитен път обаче не се използва за образуване на енергия, тъй като е:
- единствeн път за синтез на рибоза;
- основен път за натрупване на НАДФ.Н.
11.5.2. Химизъм на ПФП
Пентозофосфатният път протича в цитозола, условно се разделя на:
Ø Първи етап – окислителен – съдържа три окислителни реакции, при които се получава НАДФH и рибулозо-5-фосфат;
Ø Втори етап – синтезен - изомеразни и епимеразни реакции, които трансформират рибулозо-5-фосфат до рибозо-5-фосфат и ксилулозо-5- фосфат. Представлява серия от реакции на разкъсване на С-С-връзки и взаимопревръщания на триози, тетрози, пентози, хексози и седохептози, в резултат на което се възстановяват 5 молекули глюкозо-6-фосфат.
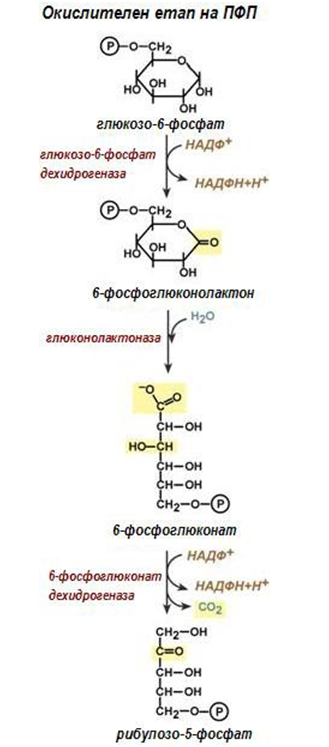
Фигура 190. Окислителен етап на ПФП
Първа реакция. ПФП започва с окислението на глюкозо-6-фосфат до 6- фосфоглюконо-δ-лактон. Приема се, че в процеса участват едновременно 6 молекули глюкозо-6-фосфат, реакцията се катализира от глюкозо-6-фосфат дехидрогеназа с кофактор НАДФ+.
Втора реакция. 6-фосфоглюконо-δ-лактонът се хидролизира от ензима 6-фосфоглюконолактоназа до 6-фосфоглюконат. Лактонът самопроизволно може да се хидролизира до свободна киселина със забележителна скорост.
Трета реакция. 6-фосфоглюконовата киселина окислително се декарбоксилира до рибулозо-5-фосфат. Ензимът фосфоглюконат дехидрогеназа (декарбоксилираща), предава отнетия водород на НАДФ+. В резултат на тази реакция хексозите се превръщат в пентози.
Вторият етап е наречен синтезен, защото единствено тук се образуват уникални метаболити с 4 и 7 въглеродни атоми, но фактически се разграждат пентози. Образуваните метаболити могат да участват в гликонеогенезата и синтезата на триглицериди.
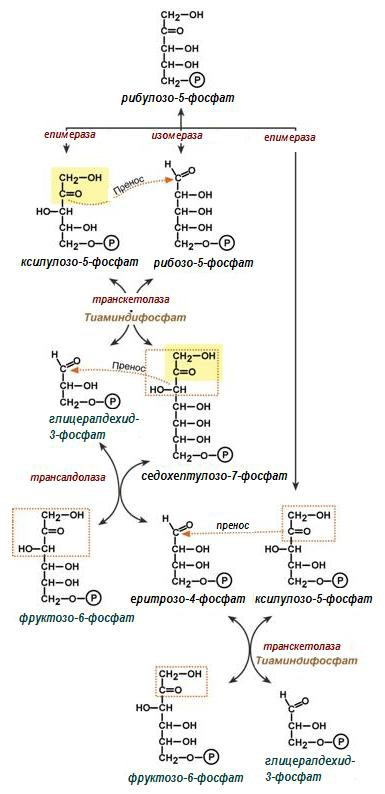
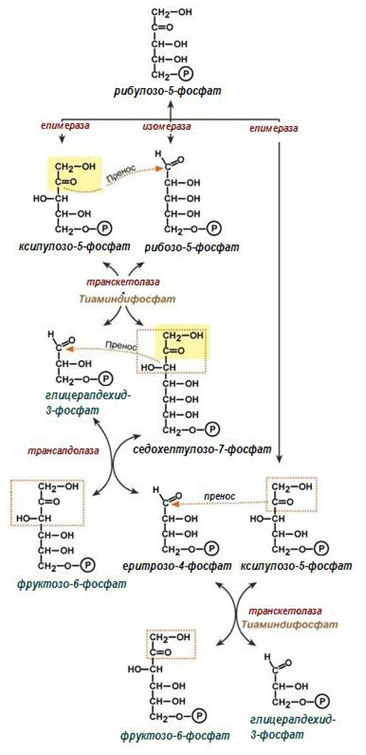
Фигура 191. Синтезен етап на ПФП
Четвърта и пета реакция. 6-те молекули рибулозо-5-фосфат се изомеризират в две молекули рибозо-5-фосфат (4) с участието на ензима рибозо-5-фосфат изомераза и в 4 молекули ксилулозо-5-фосфат (5), под действие на ензима рибулозо-5-фосфат епимераза. Рибозо-5-фосфатът е незаменим субстрат в биосинтезата на нуклеотиди, респ. НК.
Шеста реакция. Транскетолазата пренася двувъглеродни звена от ксилулозо-5-фосфата на рибозо-5-фосфата, получава се глицералдехид-3- фосфат и седохептулозо-7-фосфат.
2С5 + 2С5 ⇌ 2С7 + 2С3
Седма реакция. Трансалдолаза катализира преноса на тривъглеродни звена от седохептулозо-7-фосфата до глицералдехид-3-фосфат, при което се получава еритрозо-4-фосфат и фруктозо-6-фосфат.
2С7 + 2С3 ⇌ 2С6 + 2С4
Осма реакция. Последната трансферазна реакция, също се катализира от транскетолаза. При нея се пренасят двувъглеродни звена от другите две молекули ксилулозо-5-фосфат до еритрозо-4-фосфат с образуване на две молекули фруктозо-6-фосфат.
2С5 + 2С4 ⇌ 2С6 + 2С3
Девета и десета реакция. От останалите 2 молекули глицералдехид-3- фосфат (от реакция-8: 2С5 + 2С4 ⇌ 2С6 + 2С3) се синтезира молекула фруктозо-1,6-дифосфат, с участие на ензимите триозофосфат изомераза (9) и алдолаза (10), познати от гликолизата, но тук реакциите протичат в обратна посока. На това ниво, както и при реакции (7, 8 – фруктозо-6- фосфата), пентозофосфатния цикъл се свързва с гликолизата.
11.5.3. Значение на ПФП за метаболизма
Реакциите на биосинтеза на мастни киселини и стероиди изискват големи количества НАДФН. По тази причина, чернодробните и мастните клетки, клетките от кората на надбъбреците, тестисите и лактиращата млечна жлеза имат високи нива на ензими от ПФП. Около 30% от окислението на глюкозата в черния дроб става чрез ПФП. Еритроцитите използват големи количества НАДФН за редукция на окисления глутатион. Всички бързо пролифериращи клетки се нуждаят от високо вътреклетъчно съдържание на НАДФН. Превръщането на рибонуклеотидите в дезоксирибонуклеотиди с участие на ензима рибонуклеотид редуктаза също така изисква големи количества НАДФН.
ПФП служи като:
Ø единствен източник на пентози (преди всичко рибоза);
Ø резервоар на редуцирани форми на НАДФ, които се използват в анаболитни процеси.
Ø автотрофните организми използват еритрозо-4-фосфат за синтеза на ароматни АК.
Ø алтернативен обменен път, който при нужда доставя енергия. (Тази функция се използва от ограничен брой микроорганизми. ПФП протича в цитозола, а окислителните ензими са разположени в митохондриите, поради което пренасянето на водородните атоми от НАДФН е свързано със загуба на енергия.)
Ø икономичен обменен път по брой ензими и други химични средства.
Терминологичен речник
#гликолиза – основен, анаеробен, катаболитен път на разграждане на глюкоза в клетките, нарича се още фосфотриозен, или метаболитен път на Ембден–Майерхоф–Парнас (ЕМП). Названието «гликолиза» произхожда от гр. γλυκός, glykos — сладко и λύσης, lysis — разграждане.
#пентозофосфатният път (ПФП) - известен е и като фосфоглюконатен път, първата реакция е окисление на глюкозо-6-фосфата до глюконат. Окислението започва на ниво хексози, а не на ниво триози. ПФП окислява глюкозата и при определени условия може напълно да я разгради до СО 2 и Н2О, но се класифицира като анаболитен път, който използва 6-те въглеродни атома на глюкозата, за да образува въглехидрати с 5 въглеродни атоми.
# глюконеогенезата - синтез на глюкоза от невъглехидратни компоненти: лактат, пируват, глицерол, кетокиселини от цикъла на Кребс и други кетокиселини получавани от аминокиселини. Глюконеогенезата е път, обратен на гликолизата. Всички обратими реакции от гликолизата са част от глюконеогенеза в обратна посока на тази в гликолизата.
#глюкозо-лактатният цикъл (цикъл на Кори) - цикличен процес, обединяващ реакциите от гликонеогенезата и реакциите на анаеробната гликолиза. Глюконеогенезата протича в черния дроб, субстрат за синтезата му е лактат, постъпващ в организма от еритроцитите или мускулната тъкан.
#фотосинтеза - физико-химичен процес, при който фотосинтезиращите организми (зелените растения и цианобактерии), използват светлинната енергия за да извършват синтез на органични вещества.
#автотрофни - притежават способността да синтезират органични вещества от неорганични вещества.
#хетеротрофни - изграждат своите органични вещества от готови органични съединения, които само преработват. Хетеротрофните организми живеят за сметка на автотрофните.

