

Ключови думи: биосинтез на триацилглицероли (неутрални мазнини); фосфоацилглицероли, свободни мастни киселини; β-окисление; карнитинова совалка;
Цели:
1. Да се изясни катаболизма на мастните киселини.
2. Да се изясни функцията на карнитиновата совалка.
3. Да се изясни β-окислението като основен катаболитен път на липидите.
4. Да получат знания за биосинтезата на триацилглицеролите.
5. Да получат знания за биосинтезата на фосфоацилглицеролите.
Въведение
Понятието липид е термин от биологичната класификация на органичните съединения. Липидите са органични съединения, хетерогенна група, обединени поради своята хидрофобност. Синтезата на липидите започва от един и същи изходен метаболит ацетил-КоА.
Молекулата на липида съдържа остатъци от карбоксилни киселини, които представляват неполярните участъци на липида. Мастната киселина е свързана естерно чрез ковалентна връзка с хидроксилна група на алкохол. Причина за неразтворимостта на липидите във вода е преобладаването на неполярни химични групи в химичният им състав. Липидите са разтворими в неполярни или слабо полярни органични разтворители (етер, хлороформ, тетрахлорометан, серовъглерод CS₂, бензен и други).
Липидите се разделят на прости и комплексни. Класифицират се в няколко групи:
- Триацилглицероли (неутрални мазнини) и свободни мастни киселини;
- Фосфоацилглицероли;
- Свинголипиди;
- Стероиди;
- Восъци;
- Терпени.
Простите липиди са естери на мастни киселини с различни алкохоли. Тук спадат:
- триацилглицеролите или мазнините (естери на мастни киселини с глицерол);
- восъците - естери на мастните киселини с висши алкохоли.
При разграждане на молекулите на простите неполярни липиди (мазнините) се освобождава голямо количество енергия (около два пъти по- голяма от тази, освободена от белтъци и въглехидрати) - мазнините се явяват енергетичен източник за живите организми.
Комплексните липиди включват:
- Фосфолипиди;
- Гликолипиди (гликосфинголипиди) и други, например липопротеини;
- Сфинголипиди (основният алкохол е не глицерол, а сфингозин);
- Ацилглицеролите;
- Стероиди.
Биологичните функции на тези съединения са важни и разнообразни:
- В комплекс с белтъчните молекули изграждат мембраните на клетките и клетъчните органели и определят тяхната пропускливост;
- Изпълняват ролята на енергийни депа;
- Защитна функция – отлагат се като подкожен слой с добри термоизолационни свойства, опаковат органите и ги предпазват от механични въздействия, восъчен налеп по повърхността на листата и плодовете, липидни компоненти в бактериалната стена на бактериите – определя тяхната чувствителност към антибиотиците;
- Регулаторна функция – ейкозаноиди;
- Гликолипите са важен компонент на нервната тъкан.
13.1. Обмяна на висшите мастни киселини
13.1.1. Катаболизъм на мастните киселини
Подложените на катаболизъм в организма мастни киселини са ендогенни или постъпват в клетките от кръвта, т. е. и в двата случая са разположени в цитоплазмата и се налага транспортиране до матрикса на митохондриите, където са локализирани ензимите от катаболизма (β-окислението).
Транспортирането се предхожда от активирането им до ацил-КоА. Процесът се осигурява енергетично от АТФ. Реакцията се катализира от ацил-КоА синтетази, наричани още тиокинази. Има няколко ацил-КоА синтетази специфични за дълговерижни, средноверижни и нисковерижни мастни киселини. Разположени са в ендоплазмения ретикулум и във външната митохондрийна мембрана. Ациловият радикал се пренася върху карнитин. Ацил-карнитините се транспортират в матрикса на митохондриите, където обратно се превръщат в ацил-КоА.
1. Реакция на активиране - Мастните киселини започват своето разграждане с активиране на карбоксилната група чрез образуване натиоестери с КоА (ацил-КоА).
Преди да проникне в матрикса на митохондриите и да се окисли, мастната киселина трябва да се активира в цитозола. Това се осъществява с присъединяване на коензима А и образуване на ацил-S-КоА (фиг.229). Ацил-S-КоА е високоенергетично съединение.
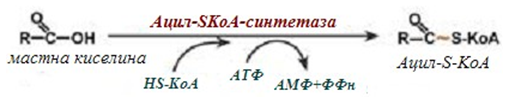
Фигура 229. Схема на активиране на мастна киселина
Необратимостта на реакцията е обусловена от хидролизата на дифосфата на две молекули фосфорна киселина.
2. Ацил-S-КоА не е способен да преминава през митохондрийната мембрана, затова съществува комплекс за пренос от карнитин. На външната мембрана на митохондрия има ензим карнитин-ацилтрансфераза I (фиг.230). Карнитин се синтезира в черния дроб и бъбреците и се транспортира в останалите органи. Във вътреутробния период и в първите години карнитин е изключително важен за организма. Карнитин е необходим за развитието на главния и гръбначния мозък.
Обезпечаването на нервната система на детския организъм, в частност на главния мозък, с енергия се осъществява за сметка на два паралелни процеса:
- карнитин-зависимото окисление на мастните киселини;
- аеробното окисление на глюкозата.
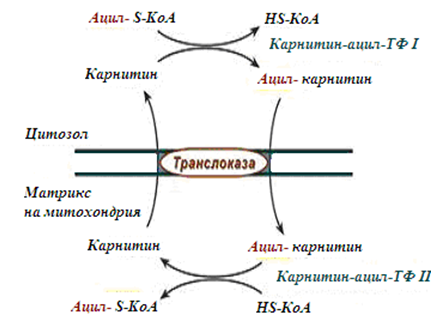
Фигура 230. Схема на карнитинова совалка
3. След свързването с карнитин мастната киселина се пренася през мембраната чрез транслоказа. На вътрешната страна на мембраната ензима карнитин-ацилтрансфераза II отново образува ацил-S-КоА който влиза в β-окислението.
4. Процес на β-окисление - състои се от 4-х реакции, повтарящи се циклично. В тях последователно протича:
- окисление (ацил-SКоА-дехидрогеназа);
- хидратация (еноил-SКоА-хидратаза);
- окисление на 3-ти С атом (хидроксиацил-SКоА-дехидрогеназа).
- В последната, трансферазна реакция от мастната киселина се откъсва ацетил-SКоА.
Към останалата (съкратена с два С) мастна киселина се присъединява HS-КоА, и тя отново се връща на първата реакция. Всичките реакции се повтарят докато в последния цикъл не се образуват два ацетил-SКоА (фиг.231).
13.1.2. Разграждане на мастни киселини с четен брой С-атоми. β- Окисление.
Главен път за разграждане на мастните киселини е β-окислението. Представлява серия от реакции с характер по-скоро на спирала, отколкото на цикъл (фиг.232). Започва с активиране на мастните киселини, при което се получават КоА-производни. Процесът е синтетазен, спрегнат с разграждане на молекула АТФ до АМФ и пирофосфат (фиг.229). Катализира се от ацил- КоА-синтетази и протича в цитозола.
Следва спираловиден процес на разграждане, в митохондриите, като на всеки оборот се откъсва по една молекула СН3СО~SКoA, която постъпва в цикъла на Кребс. При всяко преминаване по този път, въглеродната верига на мастната киселина се скъсява с два въглеродни атома, чрез отделянето на ацетатен остатък от карбоксилния край (фиг.231, 232).
β-Окислението на мастните киселини се извършва в четири реакции:
- дехидрогениране на ацил-КоА до получаване на ненаситен ацил-КоА (окислителна реакция);
- хидратиране на двойната връзка в ненаситения ацил-КоА до получаване на 3-хидроксиацил-КоА (β-хидроксиацил-КоА) - лиазна реакция;
- дехидрогениране до получаване на 3-кетоацил-КоА (β-кетоацил- КоА ) – окислителна реакция, отнема водород от и го предава по дихателната верига;
- тиолиза с КоА, при която се получава ацетил-КоА и ацил-КоА, скъсен с 2 въглеродни атома. Реакцията се катализира от ензима тиолаза. Тя е силно екзергонична и това обуславя необратимостта на целия процес.
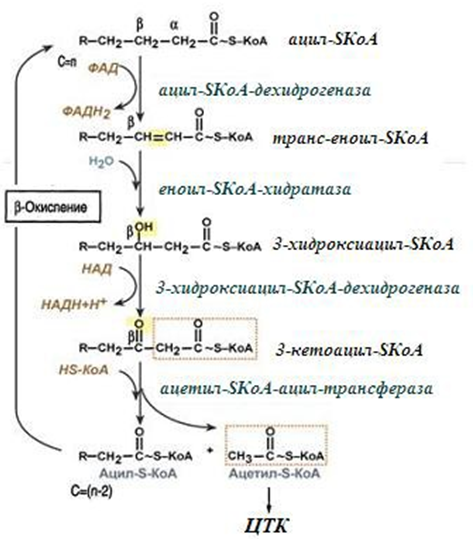
Фигура 231. Схема на β-окислението на мастните киселини
Този цикъл от реакции се повтаря докато мастните киселини с четен брой С атоми се разградят изцяло до ацетил-КоА. Полученият ацетил-СоА се отправя към ЦТК, а ацил-КоА многократно се подлага на действието на ензимите, докато изцяло се разгради до CH3CO~SКoA. При nC атоми това става за (n/2-1) цикли.
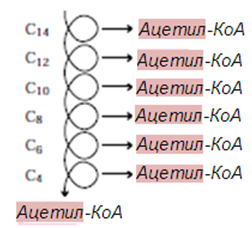
Фигура 232. β-окисление на мастни киселини, спираловиден характер
Две реакции са окислителни (фиг.231), извършва се дехидрогениране, като при едното стъпало това става с участието на ФАД (реакция 1), а при другото с НАД (реакция 3), като кофактор на съответните дехидрогенази.
Образуваните НАД.Н2 и ФАД.Н2 предават водорода и електроните по дихателните вериги до кислорода, в хода на който процес се синтезират общо 4 мола АТФ. Полученият ацетат може да се включи в ЦТК и пълно да се разгради до въглероден диоксид и вода.
β-Окислението се извършва в матрикса на митохондриите. Нарича се β- окисление, защото засяга β-атом на ациловия радикал (номер 3, фиг.233).
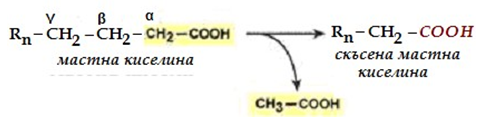
Фигура 233. Скъсяване на мастните киселини при всеки цикъл на β-окислението
В природата макар и рядко се срещат мастни киселини с нечетен брой С- атоми.
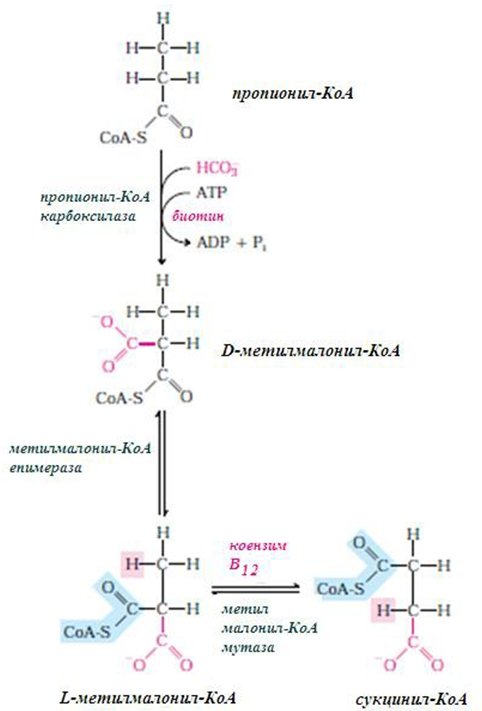
Фигура 234. Разграждане на мастните киселини с нечетен брой С-атоми, разграждат се по пътя на β-окислението, накрая се получава пропионил-КоА
Те също се разграждат по пътя на β-окислението, като накрая се образува една молекула пропионил-КоА. Пропионил-КоА може да се карбоксилира и да се получи метилмалонил-КоА, който изомеризира до сукцинил-КоА и се включва в ЦТК.
Друг път на разграждане на пропионил-КоА е β-хидроксипропионатния, застъпен в растителните организми и микроорганизмите.
13.1.3. Синтеза на мастните киселини
Синтезата на мастните киселини протича в цитоплазмата. Изходен субстрат за синтезата на мастни киселини е ацетил-КоА, който се получава в митохондриите от:
- окислително декарбоксилиране на пируват ( въглехидратната обмяна);
- разграждане на аминокиселини;
- β-окисление, което не е значителен източник на ацетил-КоА, тъй като съществува реципрочна регулация на β-окисление и синтезата на мастни киселини.
Вътрешната митохондрийна мембрана е непропусклива за ацетил-КоА, затова изнасянето на ацетилови групи през митохондрийната мембрана в цитоплазмата протича с помощта на няколко ензими и преносители в следната последователност:
1. Ацетил-КоА реагира с оксалацетат (първа реакция от цитратния цикъл) - получава се цитрат под действие на цитрат синтазата.
2. Цитратът се пренася от митохондриите в цитозола чрез трикарбоксилатен преносител.
3. В цитозола цитрат лиазата при наличие на АТФ и КоА разгражда цитрат до оксалацетат и ацетил-КоА. Ацетил-КоА се използва за синтеза на мастни киселини. Оксалацетат не може да се върне през мембраната, а се превръща в малат под действие на цитоплазмена малат дехидрогеназа.
Малатът се връща в митохондриите през дикарбоксилатен преносител и се окислява до оксалацетат, за да се затвори цикълът.
Пируват преминава в митохондриите и се декарбоксилира, превръща се в ацетил-SКоА, който влиза в ЦТК. В състояние на покой, при почивка, при наличие на излишък от енергия в клетката, на АТФ и НАДН, реакциите от ЦТК (в частност, изоцитратдехидрогеназната реакция) са блокирани. В резултат се натрупва първият метаболит от ЦТК – цитрат. По градиентната концентрация, той се премества в цитозола, разпада се и образува ацетил- SКоА, който по-нататък се използва за синтез на холестерол, мастни киселини и триацилглицероли. Оксалацетат, също се образува от цитрат, възстановява се до ябълчна киселина и се връща в митохондриите, посредством малат-аспартатната совалка; след това следва декарбоксилиране на малат до пируват от НАДФ-зависим малат-ензим.
Образуваният НАДФН се използва за синтез на мастни киселини. В цитоплазмата, с помощта на специален преносител за цитрат и задвижване на реакции се получава НАДФ.Н, необходим за синтезата на мастни киселини. Тези реакции се катализират от цитоплазмена изоцитрат дехидрогеназа и от “малат” ензим.
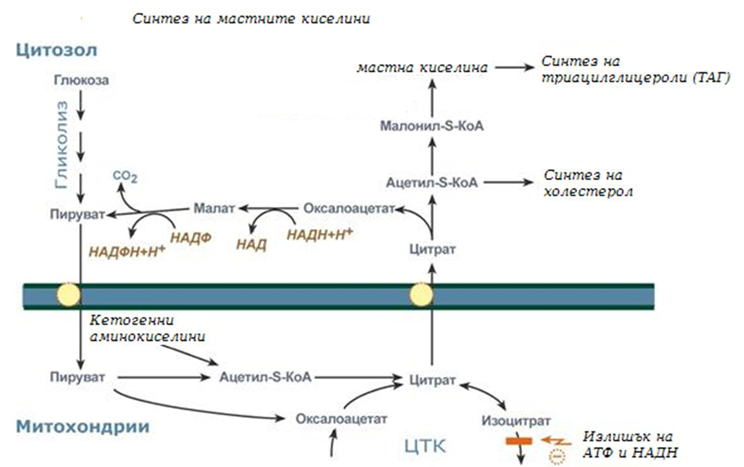
Фигура 235. Обща схема на синтез на мастни киселини и триацилглицероли
Биосинтезата на мастните киселини най-активно протича в цитозола на клетките на черния дреб, червата, мастната тъкан в състояние на покой или след хранене. Условно може да се раздели на 4 етапа:
1. Образуване на ацетил-SКоА от глюкоза, други монозахариди или кетогенни аминокиселини.
2. Пренос на ацетил-SКоА от митохондриите в цитозола:
- може от комплекса с карнитин (фиг.230), подобно на преноса на висшите мастни киселини вътре в митохондриите, но транспорта е в обратно направление;
- обикновено в състава на лимонената киселина, образувана в първата реакция на ЦТК. Постъпващият от митохондрия в цитозола цитрат се разпада от АТФ-цитрат-лиазата до оксалоацетат и ацетил-SКоА (фиг.233, 234).
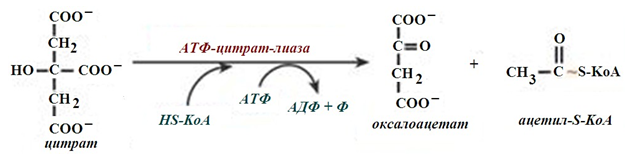
Фигура 236. Разпадане на цитрат до оксалоацетат, в цитозола от ензима АТФ-цитрат-лиаза
Оксалацетат се възстановява до малат, а след това или преминава в митохондриите чрез малат-аспартатната совалка, или от малат-ензим ("ябълчен" ензим) се декарбоксилира до пируват.
3. Образуване на малонил-SКоА от ацетил-SКоА. Карбоксилирането на ацетил-SКоА се катализира от ацетил-SКоА-карбоксилаза, мултиензимен комплекс от три ензима. Образува се малонил-SКоА от ацетил-SКоА.
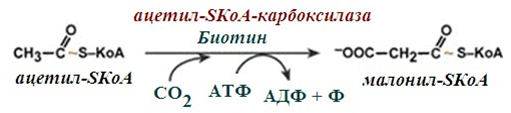
Фигура 237. Карбоксилиране на ацетил-SКоА до малонил-SКоА
4. Синтез на палмитинова киселина. Осъществява се от мултиензимен комплекс "синтаза мастни киселини" (синоним палмитатсинтаза), в чийто състав влизат 6 ензима и ацил-пренасящ белтък (АПБ).
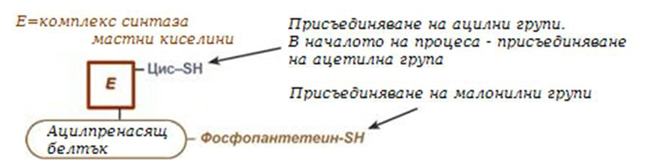
Фигура 238. Обща схема на мултиензимен комплекс "синтаза мастни киселини"
Ацил-пренасящият белтък включва производно на пантотеновата киселина – 6-фосфопантетеин (ФП), съдържащ HS-група, подобно на HS- КоА. Един от ензимите на комплекса, 3-кетоацил-синтаза, също има HS-група в състава на цистеина. Взаимодействието на тези групи обуславя началото и продължаването на биосинтезата на мастните киселини, а именно палмитинова киселина. За реакцията на синтез е необходим НАДФН.
В първите две реакции последователно се присъединява малонил-SКоА към фосфопантетеин ацил-пренасящия белтък (АПБ) и ацетил-SКоА към цистеин 3-кетоацилсинтаза.
3-Кетоацилсинтаза катализира третата реакция – пренос на ацетилна група на С2 малонил с откъсване на карбоксилна група фиг.238). След това кетогрупата влиза в реакциите на възстановяване (3-кетоацил-редуктаза), дехидратация (дехидратаза) и отново възстановяване (еноил-редуктаза), и се превръща в метиленова с образуване на наситен ацил, свързан с фосфопантетеин.
Ацилтрансферазата пренася полученият ацил на цистеин 3-кетоацил- синтаза, към фосфопантетеин и се присъединява малонил-SКоА, така цикъла се повтаря 7 пъти до образуване на остатъка на палмитиновата киселина. После тази палмитиновата киселина се откъсва от шестензимния комплекс от тиоестеразата.
Удължаване на веригата на висшите мастни киселини.
Синтезираната палмитинова киселина при необходимост постъпва в ендоплазмения ретикулум или в митохондриите. Тук с участието на малонил- S-КоА и НАДФН веригата се удължава до С18 или С20. Могат да се удължават и ненаситени мастни киселини (олеинова, линолева, линоленова) с образуване на производни ейкозанова киселина (С20). Двойната връзка в животинските клетки се въвежда не по-далеч от 9 атома на въглерода, затова ω3- и ω6-полиненаситени мастни киселини се синтезират само при съответните предшественици.
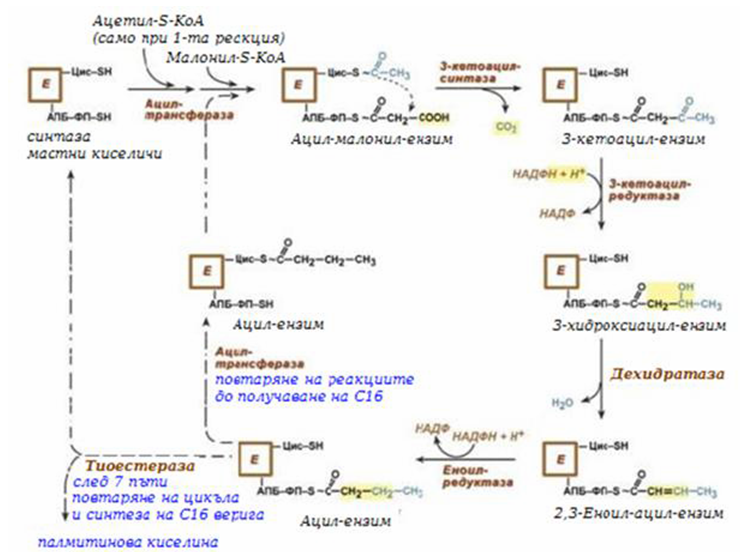
Фигура 239. Обща схема на биосинтеза на мастни киселини
Например, арахидоновата киселина се образува в клетките само при наличие на линоленова или линолева киселина. Линолевата киселина (18:2) се дехидрира до γ-линоленова (18:3) и се удължава до ейкозотриенова киселина (20:3), последната отново се дехидрира до арахидонова киселина (20:4). Така се формират мастни киселини от ω6 ред.
За образуване на мастни киселини от ω3-ред, например, тимнодонова (20:5), е необходимо наличие на α-линоленова киселина (18:3), която се дехидрира (18:4), удължава (20:4) и отново се дехидрира (20:5).
13.2. Обмяна на липидите – разграждане и биосинтез
От храносмилателния тракт липидите постъпват главно чрез лимфните пътища. В мастните клетки се складират резервните липиди под формата на триацилглицероли. При нужда те се хидролизират и мастните киселини навлизат в плазмата. Те са важен енергетичен източник за много органи.
Разграждането на липидите става според нуждите на организма. Липолиза се предизвиква под въздействието на хормоните адреналин, глюкагон, глюкокортикоиди. При състояние на напрежение, нивото на мастните киселини в кръвта се повишава 10-15 пъти и се задоволяват енергетичните нужди на организма.
За нормалното протичане на липидната обмяна е необходим инсулин. Инсулинът стимулира извънклетъчната липаза, т.е. натрупване на масти в клетките. Половите хормони регулират обмяната на липиде чрез липолиза в подкожната мастна тъкан. Естрогените имат обратен анаболен ефект – натрупване на подкожна мастна тъкан в млечните жлези и по бедрата.
Мазнини се синтезират само при наличие на енергия. Реакциите на биосинтез на липидите могат да протичат практически във всички клетки в гладкия ендиплазматичен ретикулум. Субстрат за синтеза на мазнини de novo се явява глюкозата.
Попадайки в клетката, глюкозата се превръща в гликоген, пентози и се окислява до пируват. При високи количества на постъпване на глюкоза се синтезира гликоген, но този вариант е ограничен от обема на клетката. Ето защо, глюкозата постъпва в гликолизата и се превръща в пируват, или директно, или в пентозофосфатния път. При втория вариант се образува НАДФН, който се използва за синтез на мастни киселини.
13.2.1. Разграждане на триацилглицеролите
Мазнините като вид липиди са най-разпространените липиди. Общо в мазнините са открити 50 наситени и ненаситени карбоксилни киселини. Разграждането на неутралните мазнини започва с тяхната хидролиза, която протича постепенно и се катализира от липазите. Най-напред се разграждат естерните връзки на 1-во и 3-то място, като се образуват диацилглицероли, от тях – 2 моноацилглицероли и накрая се отделя мастната киселина на втора позиция. Освободените при тази хидролиза глицерол и мастни киселини поемат свои самостоятелни пътища на катаболизъм.
Разграждане на глицерол. След фосфорилиране с АТФ и последващо окисление глицеролът се преобразува във дихидроксиацетонфосфат и се:
- включва в гликолитичната верига; или
- взема участие в глюконеогенезата.
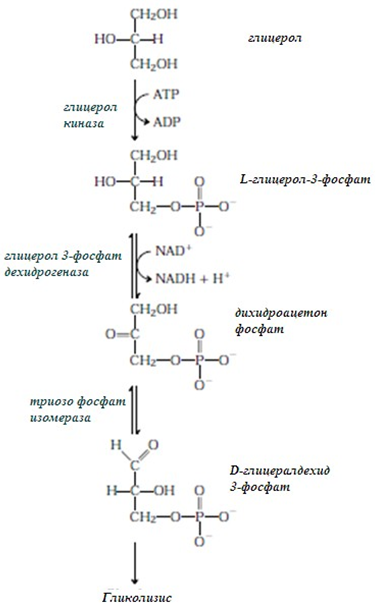
Фигура 240. Разграждане на глицерол
При разграждане на глицерол се печелят 22 mol ATP/mol.
Разграждане на мастни киселини:
- Разграждане на мастни киселини с четен брой С-атоми. β- Окисление – фиг.231;
- Разграждане на мастни киселини с нечетен брой С-атоми – фиг.234;
- Разграждане на ненаситени мастни киселини - Ненаситените мастни киселини се разграждат по пътя на β-окислението, но е необходимо наличието на специфични изомерази и епимерази, които правят възможно пълното усвояване на ненаситените мастни киселини от организмите.;
- α-Окисление на мастни киселини - β-Окислението е основен, но не единствен път за разграждане на мастните киселини. Осъществява се от два ензима – пероксидаза на мастните киселини и дехидрогеназа на дълговерижни алдехиди с коензим НАД+. Под действието на първият ензим МК се декарбоксилира и се превръща в алдехид, който се окислява от дехидрогеназата до съответната киселина.;
- ω-Окисление на мастни киселини - В някои органи мастните киселини могат да се окисляват при ω-въглеродния атом, при което се образуват α- и ω-дикарбоксилови киселини. Тяхното окисление след това продължава по пътя на β-окислението. Този път се нарича ω- Окисление на мастни киселини.
13.2.2. Биосинтеза на триацилглицероли
Изходните субстрати за синтезата на триацилглицероли (глицерол и висши мастни киселини) трябва предварително да бъдат активирани.
Глицеролът се активира чрез фосфорилиране. Под действие на глицерол киназа (фиг.241), той се превръща в глицерол-3-фосфат. Ако този ензим липсва или е неактивен, както е в мастна и мускулна тъкан, глицеролфосфат се получава от междинния метаболит на гликолитичната верига диоксиацетон фосфат.
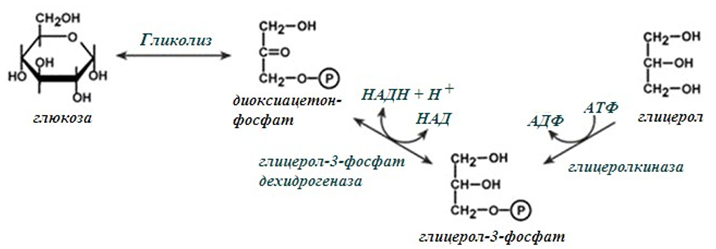
Фигура 241. Обща схема на получаване на активиран глицерол
Под действие на глицеролфосфат дехидрогеназа диоксиацетон фосфат се редуцира като водородните атоми се доставят от НАДН.
Активните форми на мастните киселини (ацил-КоА) се получават при взаимодействие на мастните киселини с КоА и АТФ (фиг.229).
Синтезата на триацилглицеролите включва четири последователни реакции:
1. Прехвърляне на ацилов радикал от ацил-КоА на първо място под действие на глицерол-3-фосфат ацилтрансфераза – получава се 1- ацилглицерол-3-фосфат;
2. Прехвърляне на втори ацилов радикал от ацил-КоА на второ място – получава се 1,2-диацилглицерол-фосфат или фосфатидат, който е предшественик за синтезата и на фосфоацилглицероли.
3. Отстраняване на фосфатната група на трето място - катализира се от фосфатидат фосфохидролаза – получава се 1,2-диацилглицерол.
4. Добавяне на трети ацилов радикал - катализира се от диацилглицерол ацилтрансфераза.
В клетките на чревната мукоза съществува и друг път, наречен моноацилглицеролен път, в който моноацилглицерол се превръща в 1,2- диацилглицерол под действие на моноацилглицерол ацилтрансфераза.
13.2.3. иосинтеза на фосфоацилглицероли
Фосфоглицеролите са главната липидна съставка на мембраните. Производни на фосфатидат, в който фосфатният остатък е естерифициран с хидроксилна група на алкохол, най-често азот-съдържащ. Мастната киселина на първо място е обикновено наситена, а на второ място ненаситена. В рамките на всеки вид може да има различни индивидуални представители в зависимост от мастните киселини на първо и второ място.
Фосфатидат е междинен метаболит в синтезата на триацилглицероли и фосфоглицероли, но не се среща в големи количества в тъканите. Двете мастни киселини образуват хидрофобен участък (“опашка”), а фосфорната киселина и алкохолът – т. нар. полярна “глава”.
В повечето фосфолипиди фосфатът е свързан с полярни съединения – серин, етаноламин (кефалини), холин (лецетини) или инозитол (инозитиди). Двете мастни киселини обикновено не са идентични. Те могат да се различават по дължината на веригата или по присъствие/отсъствие на двойни връзки. Фосфатидилхолинът (лецетин) има полярна група, създадена от холина, представлява общ мембранен липид и продуктът от неговото разграждане лизолецетин е детергент и сигнална молекула.
Лецитините са най-разпространените фосфолипиди в клетъчните мембрани. Съдържат холин - в тях се съхраняват телесните запаси на холин, който под форма на ацетилхолин е важен за предаване на нервното възбуждане и като депо на подвижни метилови групи. Важен фосфатидилхолин е дипалмитиллецитин, който предпазва от слепване вътрешните повърхности на белите дробове. Отсъствието му при някои недоносени деца води до тежко състояние (респираторен дистрес синдром).
Синтезата на фосфолипиди (ФЛ) и триацилглицероли (ТАГ) са тясно свързани. Началните етапи на реакциите на синтеза на триацилглицеролите и фосфолипидите съвпадат и протичат при наличие на глицерол и висши мастни киселини. В реакциите на биосинтез могат да се отделят следните етапи:
1. Образуване на глицерол-3-фосфат от диоксиацетонфосфат, от глюкозата или при фосфорилирането на свободен глицерол (фиг.241);
2. Биосинтез на фосфатидна киселина – изисква се наличие на глицерол-3-фосфат и мастни киселини.
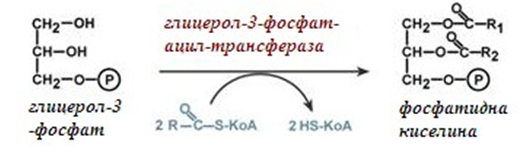
Фигура 242. Биосинтез на фосфатидна киселина
При наличие на глицерол-3-фосфат и ацил-S-КоА (активирана мастна киселина), се синтезира фосфатидна киселина.
Мастните киселини изграждащи фосфатидната киселина (фиг.242), могат да бъдат наситени или ненаситени. В зависимост от мастния състав на фосфатидната киселина, се определя синтеза, който ще протече. При наситени и мононенаситени киселини (палмитинова, стеаринова, палмитолеинова, олеинова), фосфатидната киселина се отправя за синтез на триацилглицероли. При полиненаситени мастни киселини (линоленова, арахидонова, киселини от ω3-ред), фосфатидната киселина се явява предшественик за синтез на фосфолипиди.
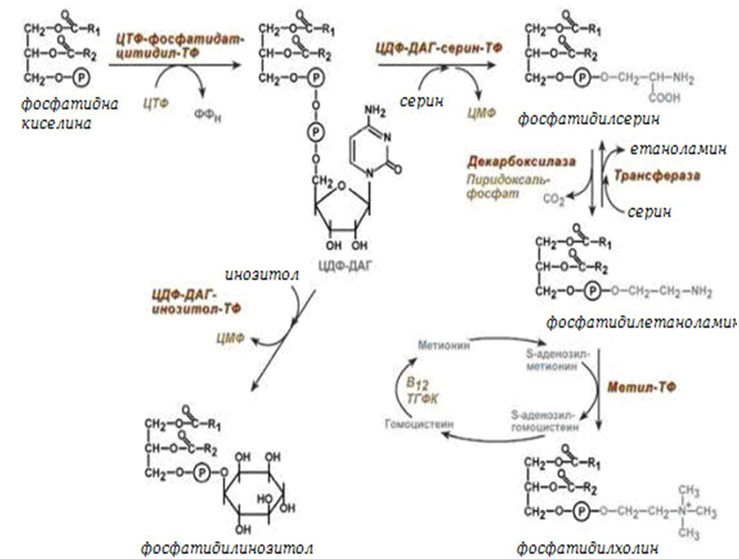
След това фосфатидната киселина се превръща – в ЦДФ-ДАГ или чрез фосфорилиране до 1,2-ДАГ. ЦДФ-ДАГ е активна форма на фосфатидната киселина способна да се превръща не само във фосфатидилинозитол, фосфатидилсерин, но и в други фосфолипиди, например в кардиолипин.
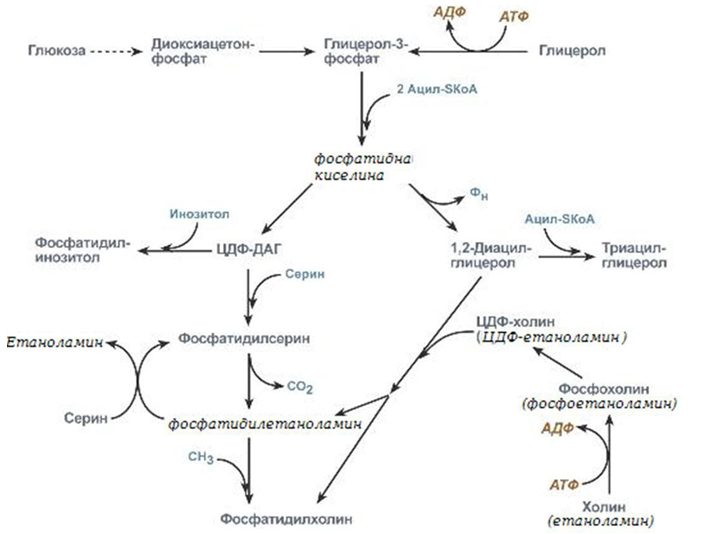
Фигура 243. Биосинтез на фосфоглицероли
Синтезираният фосфатидилетаноламин също е способен да взаимодейства със серина и обратно да образува фосфатидилсерин.
3. Синтез на триацилглицерол от 1,2-ДАГ (1,2-диацилглицерол) - след дефосфорилиране на фосфатидната киселина, образувания 1,2-ДАГ се ацилира до ТАГ.
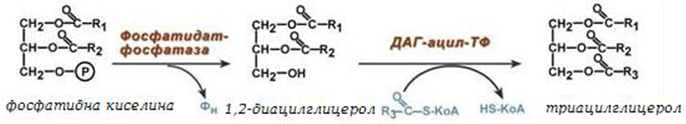
Фигура 244. Биосинтез на триацилглицероли
Синтезът на ТАГ се заключава в дефосфорилиране на фосфатидната киселина и присъединяване на ацилна група. Този процес се увеличава при следните условия:
- наличие на източник на "евтина" енергии. Например:
1) диета богата на прости въглехидрати (глюкоза, захароза) – при това концентрацията на глюкозата в кръвта след хранене се покачва и под влияние на инсулина активно преминава в синтез на мазнини в адипоцитите и в черния дроб.
2) наличие на етанол, високоенергетично съединение, при условия на нормално хранене – за пример може да служи "пивното затлъстяване". Синтезата на мазнини тук е активна в черния дроб.
- повишената концентрация на мастни киселини в кръвта, например, при усилена липолиза в мастните клетки под въздействие на вещества (фармпрепарати, кофеин и т.п.), при емоционален стрес и отсъствие (!) на мускулна активност. Синтез ТАГ при това протича в черния дроб.
- високи концентрации на инсулина и низки концентрации на глюкагон – след хранене.
След синтеза на ТАГ те се евакуират от черния дроб в другите тъкани.
4. Синтеза на фосфолипиди. Биосинтезата на фосфолипидите в сравнение със синтеза на ТАГ има съществени особености. Допълнителна активация на компонентите на ФЛ – фосфатидната киселина или холина и етаноламина.
Биосинтезата на фосфолипиди може да протече по два пътя:
- 1,2-ДАГ не се превръща в ТАГ, а се свързва с етаноламин с образуване на фосфатидилетаноламин, или с холин – и образува фосфатидилхолин.
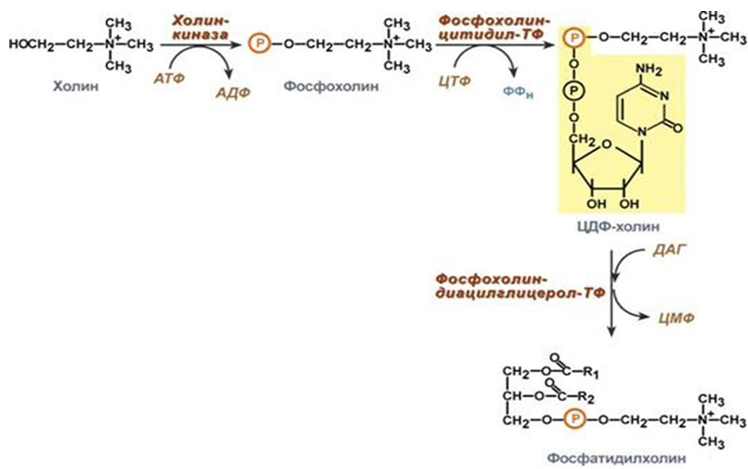
Фигура 245. Биосинтез на фосфолипиди
Холин и етаноламин се използват повторно, а не се разграждат. Активира се холина (или етаноламина)-протича чрез образуване на фосфорилирани производни с последващо присъединяване на ЦМФ. В следващата реакция фосфохолин (или фосфоетаноламин) се пренася на ДАГ.
Този начин е характерен за клетките на белите дробове, тънкото черво, но протича и в други тъкани.
Фигура 246. Биосинтез на триацилглицероли – синтез de novo
ЦДФ-ДАГ се свързва или с инозитол, или със серин с образуване на фосфатидилинозитол или фосфатидилсерин. При декарбоксилиране на фосфатидилсерина се образува фосфатидилетаноламин, който може да се превърне, във фосфатидилхолин.
Тук холин се образува от серия реакции. Активира се фосфатидната киселина с присъединяване на ЦМФ и се образува ЦДФ-ДАГ.
Терминологичен речник
#Липиди - хетерогенна група органични съединения, обединени поради своята хидрофобност. Те са неразтворими във вода, разтварят се в неполярни или слабо полярни органични разтворители (етер, хлороформ,
тетрахлорометан, серовъглерод CS₂, бензен и други).
#Прости липиди - естери на мастни киселини с различни алкохоли. Тук
спадат: триацилглицеролите или мазнините (естери на мастни киселини с глицерол); восъците - естери на мастните киселини с висши алкохоли.
#Лецитини - най-разпространените фосфолипиди в клетъчните мембрани. Съдържат холин, в тях се съхраняват запаси на холин, който под форма на ацетилхолин е важен за предаване на нервно възбуждане и като депо на подвижни метилови групи.

