

Ключови думи: клетъчна мембрана, структурата на биологичните мембрани, пермеази, унипорт, симпорт, антипорт, цитоза, пиноцитоза, фагоцитоза, ендоцитоза, екзоцитоза
Цели:
- - Да се изясни структурата на биологичните мембрани.
- - Да се изяснят основните свойства и функции на плазмената мембрана.
- - Да се изясни свойството полупропускливост.
- - Да се разгледат основните видове транспорт през мембраните.
- - Да се изяснят механизмите на клетъчен транспорт.
Въведение
Всички биологични мембрани, плазмена и вътреклетъчни мембрани на еукариотните клетки имат еднаква молекулярна организация и са изградени от два слоя липидни молекули с множество включени в тях специфични белтъци. Липидният бислой е с дебелина 4÷5 nm и е основна структура на мембраните, която създава относително непроницаема бариера за повечето водноразтворими молекули. От различните видове липиди, в плазмената мембрана, най-широко застъпени са фосфолипидите, в по-малко количество холестерол и гликолипиди.
Основните структурни особености на биологичните мембрани се определят от свойствата на липидния бислой, но техните специфични функции се осъществяват основно от белтъците. В обикновената плазмена мембрана те заемат около 50 % от нейната маса. Мембранните белтъци все още са недостатъчно проучени. Една част от тях изпълняват транспортни функции, като подпомагат преноса на определени молекули с малки размери — захари, аминокиселини, нуклеотиди и др. Втора част играят ролята на ензими, трети са рецептори на различни хормонални въздействия, а има и инертни белтъци, които участват в стабилизирането на мембранната структура.
Цитоплазмената мембрана не е просто механична преграда, която плътно обвива цитоплазмата, притежава високоизбирателна проницаемост, чрез което осъществява връзка с околната среда. В клетката проникват необходими хранителни вещества и соли, а излишните продукти на обмяната излизат от нея. Тя поддържа постоянна концентрация на цитоплазмените компоненти и отвежда ненужните на клетката продукти на обмяната.
2.1. Клетъчна мембрана - структура и функция
Клетъчната (плазмена) мембрана обгръща всяка клетка, обособява я като самостоятелна единица и определя нейната големина. Мембраната е жизненоважна, ако се разкъса се нарушава целостта на клетката и тя веднага умира. Мембраната е изградена от фосфолипиди и белтъци. Фосфолипидните молекули имат хидрофилна и хидрофобна част. Хидрофобните части на фосфолипидите, които изграждат двата слоя на мембраната, са насочени едни към други. Белтъчните структури са разположени мозаично в плътния фосфолипиден слой.
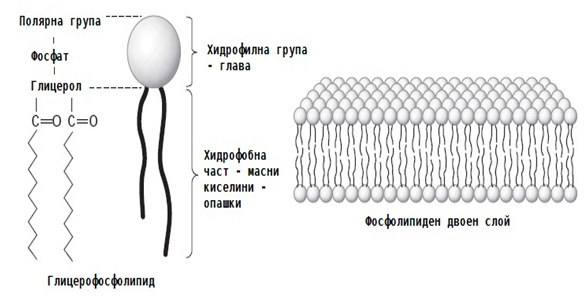
Фигура 13. Строеж на фосфолипидна молекула и фосфолипиден двоен слой
Липидните молекули в бислоя се подреждат с насочени една срещу друга хидрофобни опашки, които частично се припокриват (фиг.13). Повърхността на бислоя е съставена от хидрофилни молекули, затова бислоят е стабилен във водно обкръжение, неговата повърхност лесно се асоциира с вода. В бислоя са вградени 2 типа глобуларни белтъка:
(1) периферни – лесно могат да бъдат отделени от мембраната посредством разтвори притежаващи висока йонна сила (1 моларен NaCl) → те са свързани с повърхността на липидния бислой предимно посредством електростатични взаимодействия или чрез Н-връзки;
(2) интегрални – дълбоко потопени в липидния бислой.
Главните мембранни протеини са хидрофилни и са вградени във фосфолипидната матрица. Те могат да бъдат разделени на 3 типа:
- - изцяло потопени в бислоя – пронизват го напречно от единия до другия край, излизат от двете страни и влизат в съприкосновение с водни молекули;
- - потопени от едната или другата страна на бислоя, но не го пронизват изцяло;
- - напълно потопени в бислоя и са скрити в неговата вътрешност и не контактуват с водните молекули, обкръжаващи бислоя.
Белтъчните молекули свързани към йонните групи на фосфолипидите и водните молекули се струпват в различна степен подредени структури около външната страна на бимолекулния слой. Структурата на цитоплазмената мембрана се стабилизира от водородни и хидрофобни връзки. Катионите (Мg+2 и Са+2) също се комбинират с отрицателни заряди на фосфолипидите и помагат за стабилизирането на мембранната структура.
Всяка биологична мембрана притежава една и съща базисна молекулна организация. В резултат на проведените изследвания върху строежа на мембраните са предложени различни модели за тяхната структура, най- задоволителният и общоприет модел е течностно-мозаечен (модел на Сингер и Никълсън – 1972г., фиг.14 ). Според модела на Сингер и Никълсън, мембраната се състой от бислой липидни молекули. Липидите имат в структурата си хидрофилна (полярна) и хидрофобна (неполярна) част. Това са фосфолипиди, гликолипиди, стероли. В тях са вградени глобуларни белтъци. Типичен представител на липидите, участващи в изграждането на мембрани е фосфатидилхолин.
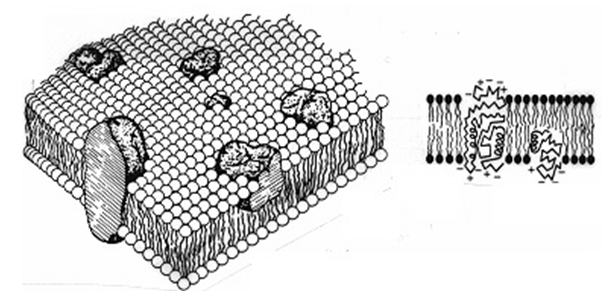
Фигура 14 . Структура на цитоплазмена мембрана (модел на Сингър и Никълсън)
Непрекъснатата част на мембраната, нейният матрикс се формира от липиден бислой, който се намира в течно състояние при обикновена за клетките температура. Мембранната повърхност не е статична структура. Липидните и белтъчните молекули могат да се преместват, много от тях се намират в непрекъснато движение. Те могат да се придвижват лесно само в плоскостта на мембраната → латерална дифузия. Възможно е и преминаване в перпендикулярна посока → напречна дифузия (флип-флоп прескок) – по-бавно движение в сравнение с горното. Фосфолипидните молекули може да се местят латерално със скорост 2 μм/sec, докато напречната дифузия на същия фосфолипид е няколко μм/h (това се обуславя от прехода на полярната глава през хидрофобната сърцевина на бислоя). Белтъците от своя страна латерално се движат по-бавно в сравнение с фосфолипидите, което се дължи на значително по-големите им размери. За тях е установена скорост от няколко μм/min. Този бислой носи множество белтъци, някои от тях са потопени частично в липидния матрикс от едната страна. Те се означават като периферни мембранни белтъци. Друга група са интегралните мембранни белтъци, които пронизват бислоя и контактуват с водната среда от двете страни на мембраната. Тези белтъци често са включени в транспорта на специфични субстанции през мембраната. По този начин мембраната има вид на мозайка от липиди и белтъци.
Мембраните в клетките се различават една от друга въпреки принципно еднаквата структура, по тип и различно съдържание на липиди в бислоя, по тип на вградени в бислоя белтъци и по съотношение на количеството белтъци спрямо липиди. Тези разлики в мембраните са свързани с техните функции. Природните мембрани притежават свойства, съгласувани с течностно-мозаечния модел, това са:
- асиметричност – външната им повърхност се различава от вътрешната. Това се обяснява с присъствието на различни белтъци върху едната или другата повърхност. Асиметрията е особено очевидна при плазмената мембрана на животинските клетки, тъй като там върху външната повърхност се намират гликолипиди, влизащи в състава на центрове за разпознаване на различни вещества;
- специфичност – природните мембрани се разпознават една друга, обуславя се от наличието върху различните мембрани, на центрове за разпознаване помежду им, като тези центрове вероятно са белтъчни. Определени мембрани в клетката могат да се сливат с някои от другите вътрешни мембрани, а други не могат, пример: митохондриите могат да се сливат (слепват) помежду си, или мехурчетата на АГ с плазмалемата, но митохондриите не са способни да се сливат с ядрото или с плазмалемата.
- избирателна пропускливост – различните клетъчни мембрани притежават различна избирателна пропускливост. Благодарение на това свойство различните компартменти на клетката могат да имат различни набори от химични компоненти и да изпълняват различна функция. Избирателната пропускливост се основава на система от преносители, вероятно с белтъчна природа, способни да разпознават определено съединение или тип съединения и да ги транспортират през мембраната, като има и носители, способни да осъществят транспорт против концентрационния градиент.
2.2. Избирателна пропускливост на мембраните
Избирателната пропускливост се отнася до качеството на биологична мембрана да пропуска само определени компоненти. Клетъчната мембрана се характеризира с това, че може да пропуска определени вещества и да не пропуска други (фиг.15). С избирателната си пропускливост регулира състава на вътрешната среда на клетката. Тъй като липидите са основен изграждащ мембраната компонент, тя е не пропусклива за полярни молекули и е лесно пропусклива за липидоразтворими вещества. Лесно преминават вещества, които са с малки и липидоразтворими молекули. Пропускливостта за йони се обяснява с наличието на специални йонни каналчета.
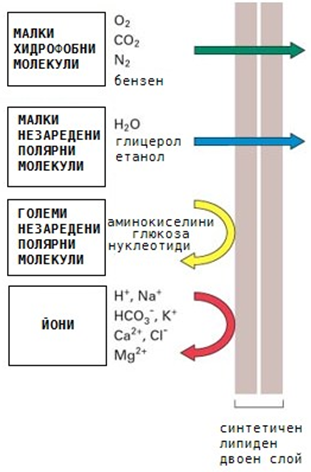
Фигура 15. Пропускливост на цитоплазмената мембрана
Малките неполярни молекули като О2, N2 и СО2 преминават през цитоплазмените мембрани без проблеми (фиг.15). Неполярни, по-големи мастноразтворими молекули, например стероидните хормони също свободно преминават през клетъчните мембрани. Свободно преминават и малките полярни незаредени молекули като Н2О и карбамид. За по-големите полярни молекули, например монозахаридите, двойният липиден слой е непропусклив. Още по-непропусклив е за йоните, дори и за най-малките като Na+.
2.3. Видове транспорт през мембраните.
2.3.1. Трансмембранен транспорт на малки молекули (през мембраната)
Едно вещество може да бъде пренесено през клетъчната мембраната по два основни механизма - придвижване със средата, в която се съдържа или чрез дифузия на веществото.
При транспортта през клетъчната мембрана е важно да се отбележи дали се използва енергия за осъществяване на транспортните процеси или не се използва. В зависимост от това транспорта бива активен и пасивен.
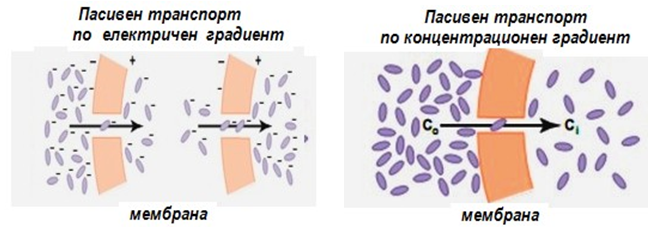
Фигура 16. Пасивен транспорт през цитоплазмената мембрана
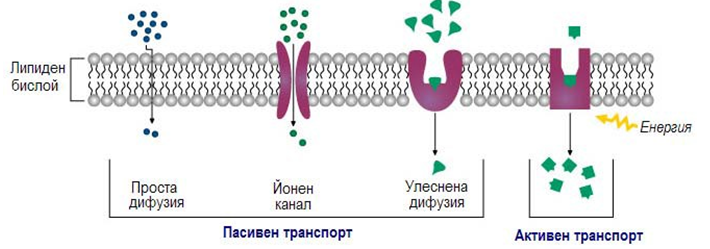
Пасивният транспорт се извършва по градиента - концентрационен, електрохимичен, хидростатичен (при разлика в хидростатичното налягане), осмотичен (при разлика в осмотичното налягане). При това свободната енергия на преминаващите молекули намалява - затова този транспорт се осъществява без разход на метаболитна енергия. Различаваме три вида пасивен транспорт:
- Дифузия:
- Обикновена (проста) дифузия;
- Улеснена дифузия;
- Дифузия през йонни канали;
- Осмоза - за да се извърши трябва да има два водни разтвора, които са разделени с клетъчна мембрана, през която могат да преминават водните молекули в посока към по-концентрирания.
- Филтрация - възниква при наличие на разлика в хидростатичното налягане от двете страни на клетъчната мембрана.
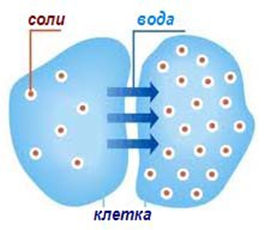
Фигура 17. Пасивен транспорт - осмоза
Обикновена (проста) дифузия. За да се осъществи пренос на дадено вещество чрез дифузия е необходимо да има химичен градиент. Ако пренасяното вещество носи и заряд, има значение и електричния градиент. При простата дифузия движението на молекулите се осъществява от място с по-висока към място с по-ниска концентрация до достигане на равновесие. Молекулите дифундират по концентрационен градиент (фиг.16). Простата дифузия е резултат от Брауновото движение на молекулите. Движеща сила е кинетичната енергия на частиците. В този случай веществото не се нуждае от мембранен преносител. При чистата дифузия потокът на веществото зависи от три фактора:
- величина на концентрационния градиент (при по-голяма концентрационна разлика ще има по-голяма чиста дифузия);
- дебелината на мембраната (при по-дебела мембрана скоростта на дифузията е по-малка);
- наличната площ на мембаната за дифузия (ако дифузионната площ е голяма, скоростта на дифузията също ще е голяма).
Малки неполярни молекули преминават през фосфолипидния слой на мембраната чрез проста дифузия, напр. О2, СО2, стероидни хормони, алкохол, анестетици и др.
Ако молекулата на пренасяното вещество няма заряд то направлението на пасивния транспорт се определя само от разликата в концентрацията на това вещество от двете страни на мембраната по концентрационен градиент (фиг.16). Ако молекулата е заредена върху посоката на нейното движение оказва влияние не само концентрационния градиент, но и общият електрически градиент през мембраната – мембранен потенциал. Двата градиента съставляват електрохимичния градиент (фиг.16). Във всяка плазмена мембрана съществува градиент на електрично поле, при което вътрешната страна на мембраната обикновено е заредена отрицателно по отношение на външната. Такъв един потенциал облекчава проникването в клетката на положително заредени йони, но възпрепятства отрицателно заредените.
Улеснена дифузия. Много мембранни транспортни белтъци дават възможност за едни или други разтворени вещества да преминават през липидния бислой посредством пасивен транспорт, тогава говорим за улеснена дифузия (фиг.18). В пасивния транспорт функционират два класа мембранни белтъци:
- белтъци – преносители;
- белтъци, формиращи трансмембранни хидрофилни канали.
Някои канали сформирани от транспортни белтъци са отворени постоянно докато други се отварят за кратко време и за тях се казва, че имат врати. Някои от тези канали се отварят в отговор на свързването на лиганди, намиращи се извън клетката посредством специфични повърхностни рецептори на клетката. Причина за отварянето на друг тип канали може да е промяна на мембранния потенциал. Трети тип канали се отварят при изменение на вътреклетъчната концентрация на някои видове йони. Например някои канали, се отварят когато се повиши концентрацията на Ca2+ йони във вътрешната страна на клетката. Много често каналите, снабдени с врати притежават самозатварящи се механизми с помощта на които каналът може бързо да се затвори независимо от това, че факторът предизвикал първоначалното му отваряне все още действа.
Улеснената дифузия се извършва по концентрационен или по електрохимичен градиент като се използва мембранен преносител за веществото, както при активния транспорт. Улеснената дифузия е по- бърза от простата дифузия поради функцията на преносителя, но с нарастването на концентрацията на транспортираното вещество преносителят се насища и скоростта на улеснената дифузия спира да расте, за разлика от простата дифузия. Транспортира се едно вещество с преносителя – унипорт (фиг.23). Преносителят търпи спонтанни конформационни промени на молекулата си, по време на които веществото се свързва с преносителя откъм средата с по-висока концентрация и се освобождава от преносителя в средата с по-ниска концентрация.
Дифузия през йонни канали. Пасивният транспорт на йоните през мембраната се причинява от дейстивето на два градиента: концентрационен (химичен) и електричен (разликата в потенциалите от двете страни на мембраната). Двата градиента действат обикновено заедно. Пасивното преминаване на йони през мембраната в резултат на едновременното действие на химичния и електричен градиент се нарича електродифузия. Сумата от двата градиента (химичен и електричен) се нарича електрохимичен градиент за даден йон, т.е. движението на йоните през мембраната се извършва по техния електрохимичен градиент. Йоните могат да преминат през мембраната по електрохимичния си градиент само чрез трансмембранни протеини - йонни каналчета или йонни преносители. Йонните каналчета не взаимодействат химически с йона - затова този транспорт е бърз. Те имат различен диаметър, електрично заредени групи иизбирателна селективност. Повечето йонни каналчета са строго селективни - пропускат само един йон (например Na+-канал). Различават се каналчета за катиони (Na+, К+, Ca2-канали) и каналчета за аниони (Cl--канали). Други са по-малко селективни и пропускат два или три йона - натриеви и калиеви, натриеви, калиеви и калциеви. Част от каналите разполагат с вратички и имат променяща се избирателна проницаемост. Отварянето (активиране) или затварянето (инактивиране) на йонните канали може да настъпи в резултат на:
- промяна в мембранния потенциал (потенциалзависими канали);
- свързване с лиганд - медиатор, хормон (лигандзависими канали);
- механична деформация на клетъчната мембрана (механозависими канали).
Фигура 18. Пасивен и активен транспорт
Активният транспорт се звършва срещу градиента и свободната енргия на преминаващите вещества се увеличава - затова е необходим разход на метаболитна енергия от АТФ. За транспортирането на определените, пренасяни вещества се използват специфични белтъци преносители, т.нар. пермеази. Активният транспорт е два вида:
- Първично активен траспорт;
- Вторично активен транспорт
Мембраната е пронизана от "мостчета" (или канали) от белтъци и именно те служат като пори, през които се осъществява регулируемия транспорт на веществата. При проучване липидното съдържание на бактерийната мембрана е установено, че тя е изградена от едни и същи липидни компоненти. Такива са фосфолипидите (главно от типа фосфатглицероли), производни на фосфолипидите, при които чрез естерификация е присъединена аминокиселина, гликолипиди и някои специфични мастни киселини (циклопропанови ненаситени и такива с разклонени въглеродни вериги).
Някои белтъци преносители работят като помпи. Помпите за разлика от преносителя са не просто малки молекули, а са комплекси, които освен преносители участват още енергодоставящи и регулаторни системи. Известни са К–Na-атефаза, протонна атефаза, Са++атефаза и анионни помпи. Преносът на протони през мембраните, в комплекса на съответната помпа участват АТФ, НАДН е получило названието протонна помпа, характерна за растителните обекти, а К–Na за животинските. Тези два вида помпи участват и имат значение за такива процеси като регулация на вътреклетъчното рН, създаването на мембранен потенциал, запасяването с енергия и нейната трансформация, осъществяване на мембранен транспорт на вещества, При активиране на протонната помпа мембранния електрохомичен потенциал на протоните нараства.
Първично активен транспорт - нуждае се от директно доставяне на метаболитна енергия под форма на АТФ. Използва се преносител - мембранен белтък, който се свързва с транспортираните йони. Преносителят притежава ензимна активност. Ензимът е АТФ-аза, която хидролизира АТФ до АДФ и неорганичен фосфат и се освобождава енергия от макроергичната фосфатна връзка на АТФ. Когато йоните се свържат с преносителя, АТФ- азата се активира и АТФ се хидролизира. Фосфатът се присъединява към транспортния протеин, който се фосфорилира и променя конформацията си, извеждайки транспортирания йон. При последващото дефосфорилиране настъпва отново конформационна промяна в преносителя и евентуално друг йон се въвежда в клетката срещу градиента.
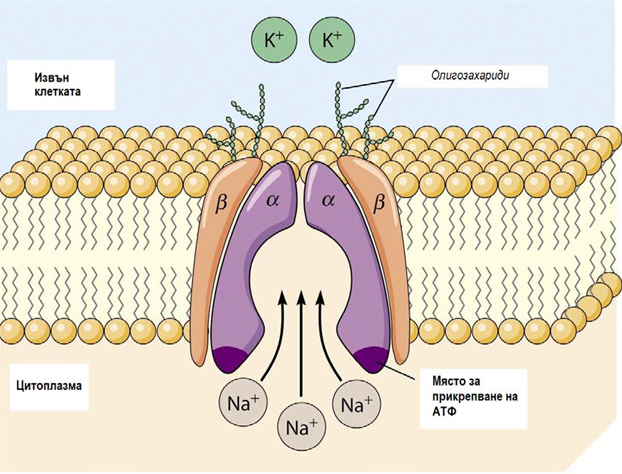
Фигура 19. Активен транспортен механизъм - Na+-K+-помпа
Типичен пример за активен транспортен механизъм е Na+-K+-АТФаза (Na+-K+-помпа) - тя присъства във всички клетъчни мембрани. Хидролизата на една молекула АТФ дава енергия за изпомпване от клетката на 3Na+ и за напомпване в клетката на 2K+ срещу техния електрохимичен градиент. Така за всеки цикъл на Na+-K+-АТФаза повече позитивни заряди се изпомпват от клетката, отколкото се вкарват вътре и се създава потенциална разлика. Вътрешната страна на мембраната се зарежда отрицателно по отношение на външната.
Вторично активен транспорт - енергията е от концентрационен градиент, създаден от протичане на първично активен транспорт, т.е. използва се индиректен внос на енергия. Ако се блокира първично активния транспорт, вторично активният не протича.
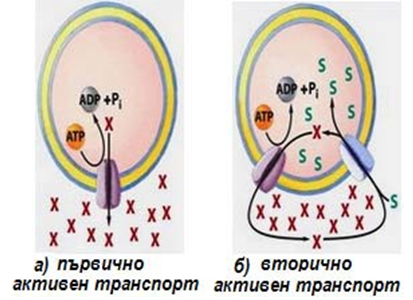
Фигура 20. Първично и вторично активен транспорт
Касае се за котранспорт с преносител, при който пренасянето обичайно на един Na+ йон е по хода на концентрационен градиент, създаван от активен транспорт (напр. Na+-K+-АТФаза), а друга молекула или йон се пренася срещу своя концентрационен или електрохимичен градиент. Na+- K+-АТФаза създава и поддържа Na+-концентрационен градиент (т.е. ниска вътреклетъчна концентрация на Na+). Котранспортът може да бъде симпорт или антипорт.
2.3.2. Везикуларен транспорт на големи молекули или частици
Везикуларният транспорт е по-особена форма на транспорт. Той не се извършва през мембраната, а чрез вгъване на самата мембрана и формиране на транспортни мехурчета. При пренос на макромолекули и по- големи частици, се извършва последователно образуване и сливане на обкръжени от мембрана мехурчета. Способността за такъв пренос се нарича цитоза.
В основата на цитозата лежи образуването на специфични вгъвания на повърхността на цитоплазмата, с помощта на която вещества намиращи се отвън се въвличат в клетката. Този процес се нарича ендоцитоза (фиг.21). Тя бива фагоцитоза, ако включеното в транспортното мехурче вещество е твърда частица и пиноцитоза, когато влюченото вещество е с течна консистенция.
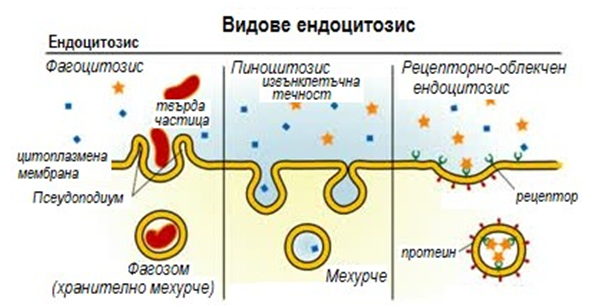
Фигура 21. Видове ендоцитозис
За да екскретира дадено вещество от клетката е необходимо клетките, които го секретират да го опаковат във вътреклетъчни мехурчета, които се сливат с плазмалемата, при което веществото се влива в извънклетъчното пространство. Този процес се нарича екзоцитоза и осъществява експорт на вещества от клетката (фиг.22).
Най – важната особеност на тези два процеса е тази, че погълнатите или секретираните вещества се локализират в мехурчетата и по този начин не се смесват с други вещества вътре в клетката.
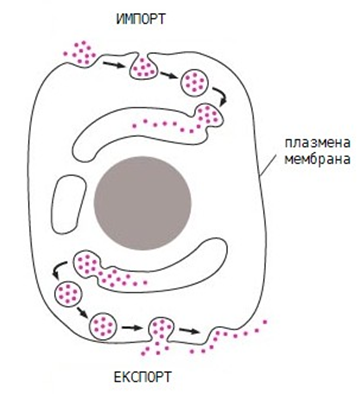
Фигура 22. Експорт и импорт на вещества извършван от цитоплазмената мембрана
2.4. Механизми на клетъчен транспорт с използване на преносител
Някои транспортни белтъци пренасят само едно разтворено вещество от едната страна на мембраната към другата такъв транспорт се нарича унипорт (унифициран транспорт). Други белтъци функционират като котранспортни системи – преносът на едно разтворено вещество зависи от едновременното или последователно пренасяне на друго вещество в същото (симпорт) или в противоположното (антипорт) направление. И в двата случая има котранспортиране (фиг. 23). При пренасяне на вещества през мембраната с използване на преносител (мембранен белтък) различаваме:
- Унипорт - преносителят пренася едно вещество;
- Котранспорт - преносителят пренася две или повече вещества.
Котранспортът може да бъде:
- Симпорт - когато веществата се пренасят в една посока.
- Антипорт - когато веществата се пренасят в противоположна посока.
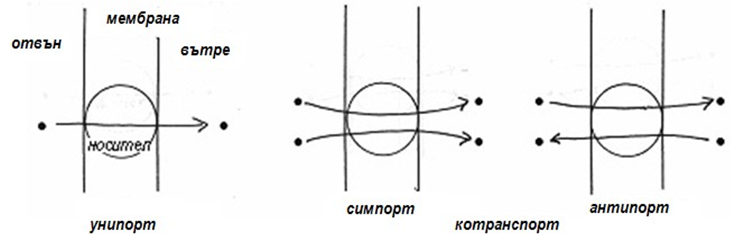
Фигура 23. Механизми на клетъчен транспорт с използване на преносител
Na+-K+-помпа в една плазмена мембрана работи на принципа на антипорта, при което Na+ се изнася извън клетката и едновременно с това K+ влизат вътре в клетката. Така например транспорт на захари вътре в много бактериални клетки става посредством насочен навътре в клетката симпорт на протони.
Терминологичен речник
Клетъчна (плазмена) мембрана - обгръща всяка клетка и я обособява като самостоятелна единица, определя нейната големина. Мембраната е жизненоважна, ако се разкъса се нарушава целостта на клетката и тя веднага умира.
Избирателна пропускливост - качество на биологична мембрана да пропуска само определени компоненти и да не пропуска други.
Пасивен транспорт - извършва се без разход на енергия.
Активен транспорт - осъществява се с разход на енергия, свързан с преминаване на вещества от място с ниска към място с по-висока концентрация. Този вид транспорт се осъществява с помощта на белтъци.
Дифузия - молекулите на веществото се движат от разтвор с по-висока концентрация, към такъв с по-ниска.
Цитоза - процеси на преминаване на вещества от околната среда в клетките или от клетките в посока към външната среда.
Екзоцитоза - процес на секреция, същността й се изразява в сливане на секреторни мехурчета (продуцирани в апарата на Голджи) с клетъчната мембрана.
Ендоцитоза - процес на преминаване на вещества от околната среда в клетките.
Фагоцитоза - клетъчната мембрана образува издавания, с помощта на които може да обхване различни твърди частици вещества.
Пиноцитоза – процес на образуване на малки мехурчета по повърхността на клетъчната мембрана, поемащи течности от околната среда.
Унипорт (унифициран транспорт) – механизъм на клетъчен транспорт с използване на преносител. Транспортни белтъци пренасят само едно разтворено вещество от едната страна на мембраната към другата.
Котранспорт - механизъм на клетъчен транспорт с използване на преносител. Преносителят пренася две или повече вещества.
Симпорт - механизъм на клетъчен транспорт с използване на преносител. Котранспорт, когато веществата се пренасят едновременно в една посока.
Антипорт - механизъм на клетъчен транспорт с използване на преносител. Котранспортна система за пренос на едно разтворено вещество, което зависи от едновременното или последователно пренасяне на друго вещество в противоположно направление.

