

Ключови думи: въглехидрати, прости и сложни въглехидрати, хомополизахариди, хетерополизахариди, гликемичен индекс, целулоза, нишесте, гликоген
Цели:
- Да се изясни химичния състав и структура на въглехидратите.
- Да се изясни класификацията на въглехидратите.
- Да се изясни биологичната роля на въглехидратите.
- Да се знаят основните представители на въглехидратите.
Въведение
Въглехидрати означава “хидрати на въглерода” (от англ. carbohydrates), по-късно се установява, че са полихидроксикарбонилни съединения. Поради сладкия вкус на много от въглехидратите се наричат още захари или захариди. Съставът им се изразява с общата формула Cm(H2O)n.
Въглехидратите (захаридите) са много разпространени в природата. В човешкия организъм са приблизително 1%, но ролята им е изключително важна. Въглехидратите са основен източник за храна. Те имат способност да се окисляват както в аеробни така и в анаеробни условия (глюкоза). Въглехидратите изграждат до 80% от сухото вещество на растенията: целулоза, нишесте, захароза, пектин и др. Глюкозата, целулозата, нишестето и гликогена принадлежат към въглехидратите. Висшите животните използват глюкозата като храна, гликогена е резервна храна, рибозата и дезоксирибозата изграждат нуклеиновите киселини. Нашето облекло е почти изцяло от целулоза, памук, лен, вискоза, ацетатни влакна и др. Мебели, къщи, хартия се получават от дърво, а то е също целулоза. Въглехидратите в буквалния смисъл обезпечават нашия живот.
Въглехидратите са основен енергиен източник на организма. Енергията, която предоставят е 17 kJ/mol (4 ккал за грам), колкото протеините, но са предпочитани, поради лесното им усвояване от организма. Те са отговорни за правилното функциониране на централната нервна система, защото са основен енергиен източник за мозъка. Той е най-големият консуматор на въглехидрати в състояние на покой. Затова недостатъчното им приемане води до състояние, познато като хипогликемия (ниска кръвна захар), което се характеризира със слабост, сънливост, раздразнителност, дори загуба на съзнание.
В организма приетите въглехидрати се разграждат до глюкоза. Тя от своя страна се разнася чрез кръвта до всички тъкани и органи. Част от нея се използва за задоволяване на енергийните потребности, а друга част се складира под формата на гликоген в мускулите и черния дроб и се използва при нужда. Когато гликогеновите депа се наситят, излишните въглехидрати се транспортират в мастните клетки, където се метаболизират в мазнини. Това е причината излишъкът от тях да ни прави дебели. За съжаление границата е тънка между достатъчния и недостатъчния им прием.
Въпреки че всеки грам въглехидрат доставя едно и също количество енергия, различните видове въглехидрати действат по различен начин върху обмяната на веществата. Това е пряко свързано с количеството на секретирания хормон инсулин след техния прием, неговите метаболитни функции и въздействие върху останалите хормони.
6.1. Свойства и биологично значение
- Енергетична функция - въглехидратите са основният и предпочитан доставчик на енергия за човешкия организъм. Те са единствения енергетичен източник за анаеробната работа на мускулите, т.е. захранват бързите мускулни влакна и главен за мозъка.;
- Резервна енергетична функция - изпълнява се от гликогена. Висшите животни използват глюкоза като енергиен източник, а гликоген като резервна храна;
- Опорно-структурна функция - вземат участие в изграждането на почти всички клетки. Въглехидратите изграждат до 80% от сухото вещество на растенията: целулоза, нишесте, глюкоза, захароза, пектин и др. Гликозаминогликаните влизат в състава на протеогликаните и участват в изграждането на съединителната тъкан (хондроитинсулфат) и т.н. Независимо, че в количествено отношение това участие е минимално, то е изключително важно.;
- Хидроосмотическа и йонрегулираща – хетерополизахаридите притежават висока хидрофилност, отрицателен заряд и по този начин задържат Н2О, йоните на Са2+, Mg2+, Na+ в междуклетъчното пространство, като обезпечават тургура на кожата и твърдост на тъканите.
- Защитно-механична функция - въглехидратите участват в изграждането на различните видове защитна слуз, която се отделя от жлезите в лигавицата на редица органи, както и в състава на ставните течности (хиалуронова киселина, гликозаминогликани).;
- Сигнална функция - олигозахаридите се намират по повърхността на клетъчните мембрани и изпълняват сигнална функция, но нямат никакво значение като хранителен продукт.
6.2. Класификация на въглехидратите
По своя строеж въглехидратите са многоатомни алкохоли с алдехидна или кетонна група (полихидроксиалдехиди и полихидроксикетони). Най- известните въглехидрати (скорбяла, глюкоза и гликоген) притежават обща емпирична формула (СН2О)6. Други представители на въглехидратите, включват в състава на молекулите си атоми на азота, сярата и фосфора.
Класификацията на въглехидратите се изгражда въз основата на състава и структурата на молекулите, според което се разделят се на две основни групи: прости и сложни.
Номенклатурата на IUPAC е утвърдила окончанието –оза в наименованията на по-голяма част от въглехидратите.
6.2.1. Прости въглехидрати
Съгласно съвременната класификация, простите захариди се подразделят на 3 групи: монозахариди, олигозахариди и полизахариди.

Фигура 60. Класификация на въглехидратите
Монозахариди (монози)
В молекулата на монозахаридите въглеродните атоми са свързани в права (нормална) верига и при един С-атом може да има само една –ОН- група. Класификацията на монозахаридите се основава на съчетаването на два признака:
- вида на функционалната карбонилна група;
- броя на С-атоми в молекулата.
Според първия признак монозахаридите се делят на алдози (съдържат една алдехидна група, -СНО) и кетози (съдържат една кетонна група, С=О), а според втория се разделят на – триози, тетрози, пентози, хексози и т.н. Монозахаридите са бифункционални съединения и се различават според типа на карбонилната група (алдози и кетози), броя на въглеродните атоми в молекулата, брой и пространствено разположение на асиметричните (хирални) въглеродни атоми.
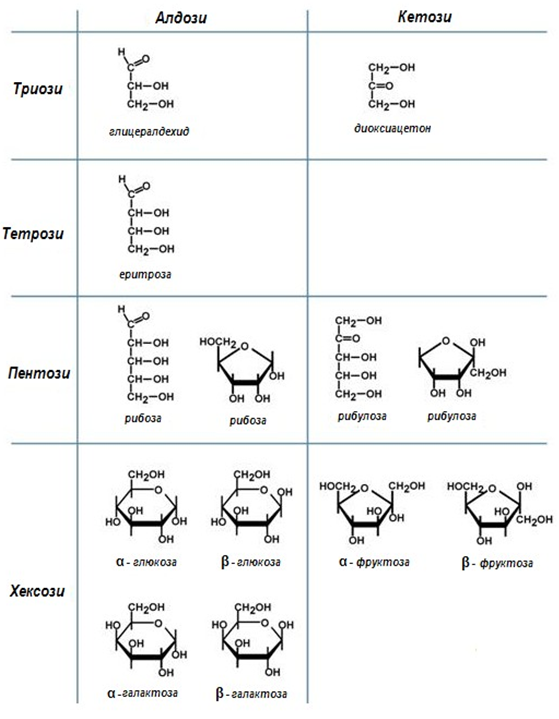
Фигура 61. Класификация на монозахаридите
Изомерни форми на монозахаридите се получават въз основа на:
- конформацията на асиметричните въглеродни атоми, стереоизомери: L- и D- форми, формиращи L- и D- стеричен ред (енантиомери);
- конформацията на първия въглероден атом на въглерода – α- и β- форми (аномери);
- по конфигурацията на хидроксилната група на един от хиралните водородни атоми в молекулата (епимери);
- алдо- и кетоизомерия, присъствието на алдехидна или кетонна група – алдози или кетози.
Алдози. В техните молекули алдехидната група естествено е при краен С-атом, а при останалите С-атоми има по една –ОН-група. Според броя на въглеродните атоми биват: алдотетрози, алдопентози, алдохексози и т.н. По-голямо значение имат алдопентозите и алдохексозите, много от които са природни продукти.
Най-простият представител на алдозите е глицериналдехида (2,3- дихидроксипропанал, глицералдехид). Той притежава един асиметричен (хирален) С-атом и следователно има два енантиомера:
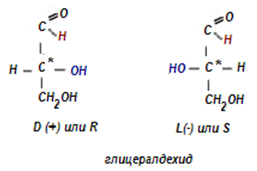
Фигура 62. Енантиомери на глицералдехида
Условно е било прието с D(+) да се означава този енантиомер, в проекционната формула на който хидроксилната група е отдясно (фиг.62), когато СНО-групата е отгоре. Другият енантиомер е означен като L(-). Не съществува ясна зависимост между структурата на молекулата и посоката на въртене. Така че дадено съединение може да бъде D(+), D(-), L(+) или L(-). Смес от равни количества от D и L форми няма оптична активност, тъй като активностите на двата изомера се неутрализират. Такава смес се нарича рацемична. Конфигурацията на D-енантиомера съответства на R- конфигурация, а тази на L-енантиомера – на S-конфигурацията, по R, S- системата на Кан-Инголд-Прелог.
D, L-системата се основава на следната постановка: монозахариди се разглеждат като произлезли от D-глицериналдехида чрез удължаване на въглеродната верига откъм СНО-групата, се обединяват в т.нар.D-стеричен ред енантиомери (стереоизомери), по аналогичен начин се регламентира формирането и на L-стеричния ред. По-висшите от глицериналдехида алдози притежават нарастващ брой структурно различни асиметрични С- атоми и имат 2n хирални стереоизомера. Те се систематизират съответно в D, L-стерични редове според конфигурацията на най-отдалечения от алдехидната група асиметричен С-атом. Енантиомерите на дадената алдоза имат еднакви наименования окончаващи на –оза, чрез които се означава конфигурацията на всеки от тях: D- или R , съотв. L- или S. При алдохексозите има четири хирални центра, познати са 16 стереоизомера (D- и L-): глюкоза, маноза, идоза, галактоза, алтроза, алоза, гулоза и талоза.
Въглехидратите са многоатомни алкохоли с алдехидна или кетонна група (полихидроксиалдехиди и полихидроксикетони). Алкохолите могат да реагират с алдехиди и кетони, при което се получават съответно хемиацетали и хемикетали (фиг.63). По подобен начин в монозахаридите с повече от четири въглеродни атома хидроксилните групи могат да реагират с алдехидната (или кето-) групата. Водороден атом от една от хидроксилните групи преминава към карбонилната група.Така се получават полуацетални и полукетални производни с циклична пръстенна структура.
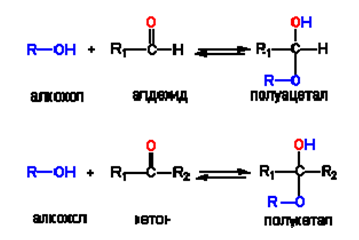
Фигура 63. Получаване на полуацетал или полукетал
Преминаването на карбонилната в полуацетална или полукетална форми е обратимо и се извършва спонтанно във воден разтвор. Обратимото преминаване на монозахаридите от карбонилна в полуацетална или полукетална форма има голямо значение в биохимията. При някои биохимични реакции монозахаридите се включват със своите карбонилни, а при други - със своите полуацетатни или полукетални форми. При циклизирането се създава нов хирален център и съответно на това два полуацетални изомера. Новообразуваната полуацетална хидроксилна група при този център се отличава с по-висока химична реактивност и с по-особени биохимични отнасяния. В бихимията за нея се предпочита названието гликозидна хидроксилна група.
Пръстените биват шест атомни “пиранозни” (пет въглеродни и един кислороден хетероатом) или пет атомни “фуранозни” (четири въглеродни и един кислороден хетероатом, фиг.64).
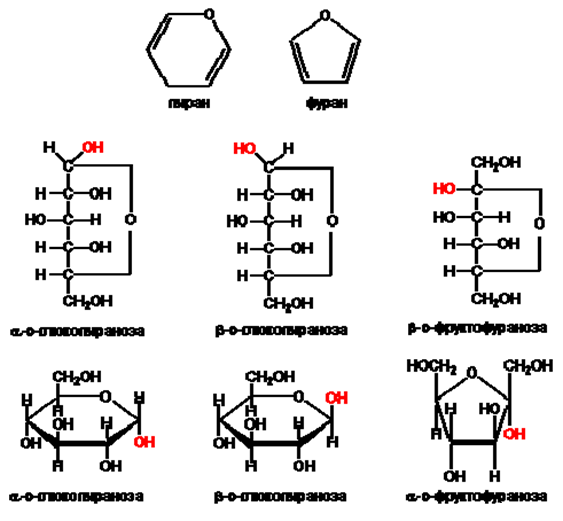
Фигура 64. Полуацетални и полукетални форми на хексози, Фишерови и Хейуъртови проекции.
Първите се наричат “пиранозни” (от структурата на хетеропръстенното съединение пиран), а другите се наричат “фуранозни” (от структурата на хетеропръстенното съединение “фуран”).
Вместо плоскостните Фишерови проекции за изписване на цикличните форми на монозахаридите се използват обикновено пространствените структури, известни като Хейуъртови проекции, или формули. Хидроксилните групи, които са отдясно във формулите на Фишер са отдолу при формулите на Хауърт (мнемонично правило: дясно – долу).
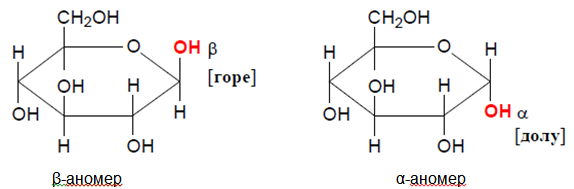
Фигура 65. Циклични формули на Хауърд, α-аномер и β-аномер на глюкозата
При формулите на Хауърд трябва да се знае, че винаги в шестчленния цикъл кислородния атом се намира в горния десен ъгъл. Лежащите извън пръстена въглеродни атоми, например шестият въглероден атом при D- глюкопиранозата, също се изписват над плоскостта на пръстена. В разтвор D-глюкозата е над 99% под форма на D-глюкопираноза, а при фруктозата преобладава D-фруктофуранозната форма.
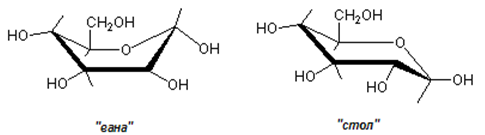
В действителност атомите (въглеродни и кислороден), които образуват пиранозните или фуранозни структури на захарите не лежат напълно в една плоскост. При шестатомния пиранозен пръстен са възможни две пространствени конфигурации на пръстена, обозначени като “вана” и “стол” (фиг.66). Енергетично предпочитаната конфигурация е тази на “стол”.
Фигура 66. Пространствени конфигурации „вана“ и “стол”
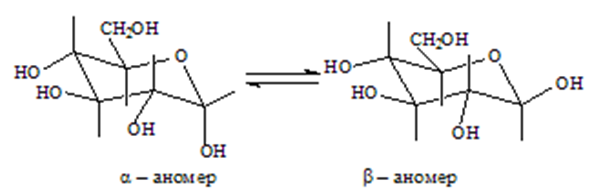
Пространственото разположение на хидроксилните групи при хиралните въглеродни атоми се изразява най-ясно с конформационните форми “стол”:
Фигура 67. Конформационна форма “стол”, α-аномер и β-аномер на глюкозата
По-стабилна е молекулата на β-изомера, тъй като всички хидроксилни групи се намират в по-изгодното екваториално положение. Ето защо целулозата е най-разпространения въглехидрат в природата (съставена от β-D-глюкозни остатъци).
Кетози. При кетозите карбонилния С-атом (С=О) е най-често втория въглероден атом, според броя на въглеродните атоми също се разделят на кетотетрози, кетопентози, кетохексози и т.н (фиг.61). Както при алдозите и тук стереоизомерите се причисляват към D, L-стерични редове (енантиомерия).
Представители на монозахаридите
D-глюкоза (гроздена или гроздова захар) – естествени източници на глюкоза са плодовете, най-високо съдържание има в гроздето (затова се нарича още гроздова захар). Усвоява се най-бързо от всички въглехидрати, като този процес зависи от наличието на хормона инсулин. В организма всички приети въглехидрати се метаболизират до глюкоза. У здрав човек, в кръвта и в урината, тя е около 0.1%, а при болен от диабет – 12%. Концентрацията на захар в организма се регулира от хормон, наречен инсулин. В чист вид глюкозата се добива чрез ензимна хидролиза на нишесте (царевица, ориз, картофи, пшеница) и се използва главно в сладкарската промишленост. Производно на D-глюкозата е витамин С, за синтеза на вит.С се излиза от D-глюкоза. Той се намира в много растения – шипки, цитруси, пиперки, магданоз и др.
D-маноза – намира се в портокаловите кори и в охлювите.
D-фруктоза (овощна захар) – или плодова захар се съдържа в сладките плодове и меда (около 8%) под формата на инвертна захар. Усвоява се два пъти по-бавно от глюкозата, като не изисква инсулин, затова е известна и като захар за диабетици. Фруктозата лесно се конвертира в мазнини. В мускулите не се използва директно, складира се в черния дроб под формата на гликоген. Средно там се съхраняват около 100 грама въглехидрати и когато депата са запълнени, излишната фруктоза заминава директно в мастните клетки. Качеството й да не покачва инсулина не е за пренебрегване и употребявана разумно, тя може да намери своето място във всяка една диета. Още повече, основните й източници – плодове и мед, съдържат и други ценни съставки.
Галактоза - постъпва в организма главно като съставна част на дизахарида лактоза, който се съдържа в млякото и някои млечни продукти. Взима участие в енергийната обмяна след като предварително се превърне в глюкоза.
Производни на монозахаридите
Уронови киселини – това са карбоксилни производни, при които най- отдалечената от карбонилната група хидроксилна група е окислена до карбоксилова (уронови киселини). Влизат в състава на протеогликаните.
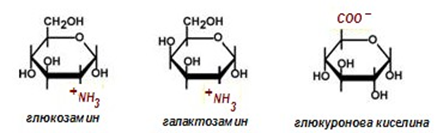
Фигура 68. Производни на монозахаридите
Аминозахари – една от хидроксилните групи (най-често тази на второ място) е заместена с аминогрупа (фиг.68).
Гликозиди – съединения, които се образуват по пътя на кондензация на монозахарид (свободен или свързан в полизахарид), с хидроксилната група на друго съединение, което може да бъде друг монозахарид или метанол, глицерол, стерол, фенол.
Дезоксизахари - една от хидроксилните групи липсва (дезоксирибоза).
Дизахариди
Дизахаридите са изградени от по два монозахарида. Установено е, че свързването на две молекули монозахарид в молекула дизахарид става чрез обезводняване между две ОН-групи на монозахаридите, поне едната от които е гликозидна (гликозиден е този С-атом, който преди това е бил в състава на карбонилна група). Гликозидното свързване между монозахаридите, може да бъде трехалозов, малтозов, гентиобиозов и целобиозов тип свързване (фиг.69).
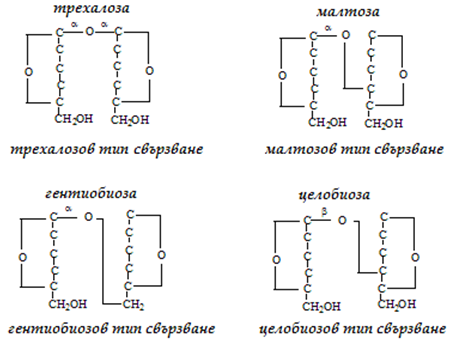
Фигура 69. Видове дизахариди, според типа на гликозидно свързване
При използване на цикличните проекционни формули на Хауърд имаме:
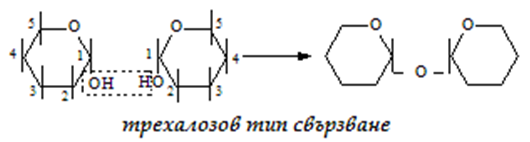
а) трехалозов - 1-1 тип свързване, с участие на 2 гликозидни ОН-групи:
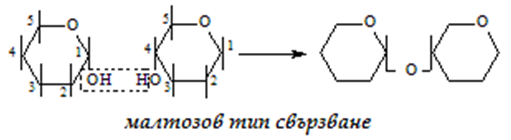
б) малтозов - 1-4 тип свързване, с участие на 1 гликозидна и 1 алкохолна ОН-групи:
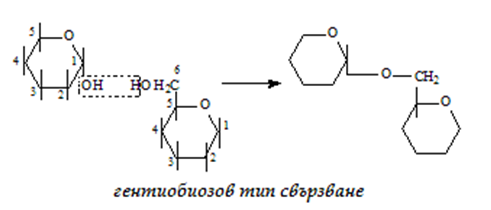
в) гентиобиозов - 1-6 тип свързване, с участие на 1 гликозидна и 1 алкохолна ОН-групи:
г) целобиозов - 1-4 тип свързване, но гликозидната ОН-група е в β-положение:
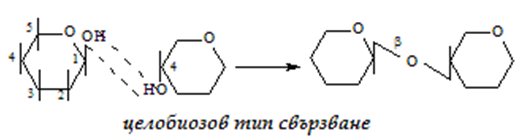
Тези няколко типа свързване между монозахаридите са в основата на образуване на молекулите не само на дизахаридите, но и на полизахаридите като напр. нишесте и целулоза.
Представители на дизахаридите
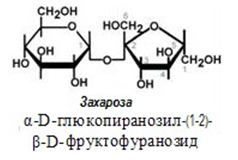
Захароза - обикновена захар, получава се от захарно цвекло или от захарна тръстика. Изградена от една молекула глюкоза и една фруктоза, тя е α-D-глюкопиранозил-β-D-фруктофуранозид. Молекулата и е образувана чрез трехалозов тип 1-2 свързване на два гликозидни С-атома, т.е. две гликозидни ОН-групи - на α-D-глюкопираноза и β-D- фруктофураноза:
Малтоза - продукт от хидролизата на скорбялата и гликогена, които се разпадат по време на храносмилането, съставена е от две молекули глюкоза, свързани с α 1,4 – гликозидна връзка. Не се среща самостоятелно в храната. При разграждането на нишестето се образуват известни количества малтоза, които след това се хидролизират до глюкоза.
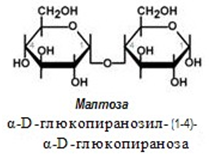
Малтоза (малцова захар) се получава от нишесте с помощта на ензима диастаза, който се намира в малца (прорасли и изсушени ечемичени зърна). Същото действие върху нишестето има и птиалинът на слюнката. Малтозата е по-малко сладка от захарозата.
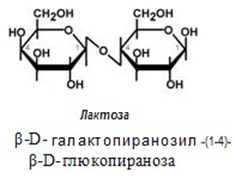
Лактоза - млечна захар, съдържаща се в млякото и някои млечни продукти. Изградена е от една молекула глюкоза и една галактоза. Тя има голямо значение за кърмачетата и по-малко за възрастните. Голяма част от хората развиват различна степен на непоносимост към лактозата, тъй като имат занижени нива на ензима лактаза, който я разгражда. Такива индивиди не могат безпроблемно да консумират прясно мляко и неферментирали млечни продукти, но в различна степен се справят с приема на кисело мляко, сирене, извара.
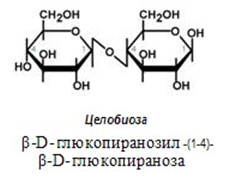
Целобиоза – междинен продукт от хидролизата на целулозата в тънкото черво. При здрава микрофолора ¾ от постъпващата целулоза се разпада до свободна глюкоза, която или се употребява от самите микроорганизми или се абсорбира в кръвта.
6.2.2.Сложни въглехидрати (Полизахариди)
Сложните въглехидрати (незахароподобни полизахариди) са изградени от стотици или хиляди монозахарида (мол. маса средно 100 000 Da). Полизахаридите са полимерни въглехидрати, образувани от повтарящи се моно- или дизахариди, свързани заедно чрез гликозидна връзка. В зависимост от състава на изграждащите ги монозахариди се разделят на хомополизахариди (еднакви монозахариди) и хетерополизахариди (различни монозахариди). Те са линейни неразклонени или разклонени в различна степен.
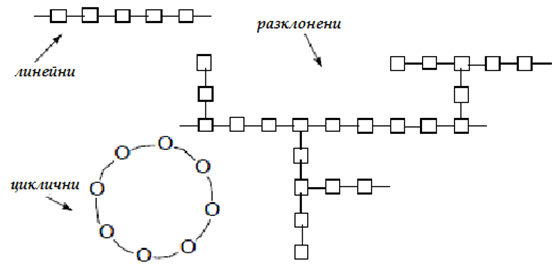
Фигура 70. Видове полизахариди, според типа на мономерите и строежа на веригата
Представители на полизахаридите
Полизахаридите изпълняват структурна и резевна функция в организма. Представители на хомополизахаридите са скорбялата, гликогена и целулозата, а на хетерополизахаридите - хиалуроновата киселина, хондроитинсулфат.
Хомополизахариди
Биополимерите на глюкозата (напр. целулоза, нишесте) се означават с общото название глюкозани. Декстраните са силно разклонени глюкозани, които се образуват при ферментация на захароза, причинена от бактерии; техни водни разтвори се използват като заместители на кръвната плазма. Прилагат се също във фармацевтичната технология.
Нишесте (скорбяла) – хомополимер на α-D-глюкозата, основен въглехидрат доставящ енергия за нашия организъм, в храносмилателния тракт се хидролизира до глюкоза. От 100 грама нишесте се получава 105 грама глюкоза, поради включването на молекула вода при хидролизата на връзките. Молекулите на нишестето са изградени от α-D-глюкопиранозни остатъци, свързани чрез поликондензация от малтозов (1-4 свързване) и гентиобиозов тип (1-6 свързване).
Нишестето е нееднородно и се състои от две вещества: амилоза (15- 20%), и амилопектин (80-85%). Амилозата е разтворима във вода, а амилопектина не се разтваря а само набъбва. Основните хранителни продукти съдържащи нишесте са зърнените култури – ечемик, пшеница, елда, ориз, овес, просо, ръж, царевица и др., както и получените от тях брашна, т.е. в хлебните продукти. Картофите и бобовите храни също са богати на нишесте.
Амилоза. Молекулата на амилозата е изградена чрез 1-4 свързване на α- D-гликозидни остатъци. Тя съдържа около 200– 1000 подобни остатъци и има спираловидна или т.нар.helix-структура в права линия:
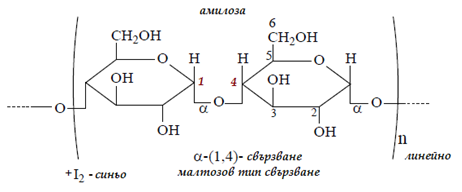
Фигура 71. Структура на амилозата, малтозов тип свързване, 1-4 свързване на α-D-гликозидни остатъци
При хидролиза амилозата изцяло се превръща в малтоза. Молекулната маса на амилозата варира в границите 30 000 - 220 000 Da. Тъй като молекулите на амилозата са макромолекули, тя може да се разглежда като природен полимер на глюкозата.
Амилопектин. Молекулата на амилопектина е разклонена, тъй като освен 1-4 свързване се осъществява и 1-6 свързване (гентиобиозов тип). Средно на 20-30 линейни звена се извършва по едно 1-6 свързване. Амилопектина съдържа ~2500 глюкозни остатъка:
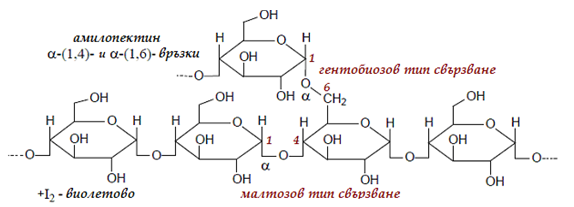
Фигура 72. Структура на амилопектина, малтозов и гентиобиозов тип свързване, 1-4 и 1-6 свързване на α-D-гликозидни остатъци
Мястото на 1-6 свързването се нарича точка на разклонение. Въпреки че разклонените вериги са по-къси от линейните вериги на амилозата, поради големия брой на разклоненията молекулната маса на амилопектина е по- голяма от тази на амилозата. Частичната хидролиза на амилопектина дава декстрини – достатъчно едри молекули, съдържащи 8-12 глюкозни остатъци. По-нататъшната хидролиза дава дизахариди – главно малтоза. При пълната хидролиза се получава α-D-глюкоза.
Нишестето е бяло микрокристално вещество, неразтворимо в студена вода, без вкус и мирис. В гореща вода набухва, образувайки колоиден разтвор. При реакция с йодов разтвор се оцветява в синьо, което се използва като индикатор за присъствието му. В стомашния тракт на човека и животните нишестето се разтваря, подлагайки се на хидролиза под въздействие на ензима амилаза и се превръща в глюкоза, която се усвоява от организма. Поради това то е един от основните енергийни източници в ежедневното хранене на човека, тъй като се съдържа в изобилие в хляба, картофите и зърнените храни. Основна съставка на брашното (75-80%), картофите (до 25%) и др.
Нишестето намира приложения в хранителната, хартиената и фармацевтичната промишленост за получаване на етилов алкохол, глюкоза, хартия, текстил, лепила и др. В промишлеността превръщането на нишестето в глюкоза (процес на озахаряване) се извършва чрез неколкочасовото му варене в разредена сярна киселина. Така нареченото модифицирано нишесте се използва за сгъстяване на замразени храни. Ако е необходимо да се получи чиста глюкоза, варенето на нишестето продължава, докато се постигне по-пълното му превръщане в глюкоза. При нагряването на сухо нишесте до 200-250° се извършва неговото частично разлагане и се получава смес от по-малко сложни от нишестето полизахариди (декстрин и др.). Намира приложение и при получаването на витамини и антибиотици. Получените при частичната хидролиза на нишестето декстрини се използват за приготвяне на декстринови лепила. Колата или нишестеният клей се използват за колосване на памучни тъкани.
Гликоген - резервен захарид, съставна част на нашия организъм, складова форма на глюкозата. Всяка клетка съдържа малки количества гликоген, но големи натрупвания има само на две места в организма – в мускулната тъкан и в черния дроб. Гликогена (”животинско нишесте”), се образува в черния дроб и мускулите на животните и играе важна роля в метаболизма на въглехидратите. Молекулната му структура е аналогична с тази на амилопектина, но е по-разклонена, средно на всеки 8-10 глюкозни остатъка има по едно 1-6 свързване и дължината на разклоненията са по- къси 11-18 глюкозни остатъци. Поради тези особености, той е значително по- компактен, мускулите съдържат много повече гликоген (300-600 грама), в сравнение с тежащия около килограм черен дроб (60-100 грама). Мускулния гликоген доставя глюкоза само за мускулната работа, а чернодробния снабдява с глюкоза всички тъкани и органи.
Целулоза – най-разпространеното съединение в биосферата, приблизително половината въглерод на планетата влиза в нейния състав. Името и произлиза от cellula – клетка, съставна част на клетъчните стени на растенията. Тя има линейна структура на молекулата, изградена от β-D- глюкопиранозни остатъци, свързани с β-1,4-гликозидна връзка (фиг. 73). Средният брой на глюкозидните остатъци в молекулата е 600-6000, а молекулната маса вече е доста голяма – от 100000 до 1000000. За разлика от разгледаните полизахариди, има нишковидна структура и е абсолютно неразтворима във вода. Установено е, че линейните молекули са свързани “сноповидно” (фибрилно) във влакна чрез междумолекулни водородни връзки. Това придава значителна здравина на целулозните влакна.
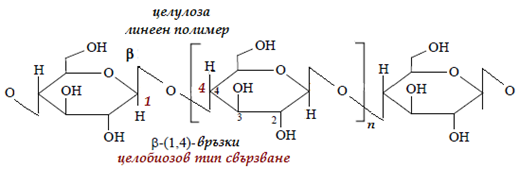
Фигура 73. Структура на целулозата, целобиозов тип свързване, β-1-4 гликозидна връзка
При частична хидролиза на целулозата се получават олигозахариди – главно целобиоза, а по-нататък – β-D-глюкоза. Целулозата е полимер на глюкозата, но поради разлика в строежа на връзките между отделните глюкозни молекули, е напълно несмилаема и няма никаква енергетична стойност. Причината за това е следната: глюкозните звена в молекулата и са в β-аномерна форма на конфигурацията “стол”, което е най-стабилната структура на глюкопиранозата поради екваториалното разположение на всички ОН-групи. Това определя и разликата в стабилността и свойствата и в сравнение с нишестето, в чиято молекула глюкозните остатъци са в α- аномерна форма. Нейната роля е да подобрява перисталтиката на стомаха и червата, да стимулира отделянето на храносмилателни сокове и да създава чувство за ситост. Заедно с растителната храна ние поемаме значителни количества целулоза. Освен същинската целулоза са познати още хидратна целулоза, хемицелулоза, пектин, хитин и др.
Хетерополизахариди
Повечето хетерополизахариди се характеризират с повтарящи се дизахаридни остатъци. Тези дизахариди включват в състава си уронова киселина и аминозахар. Дублирайки се образуват олиго- и полизахариди – гликани. Използват се синонимите – кисели хетерополизахариди, гликозаминогликани (производни на глюкозата, съдържат аминогрупа). Основни представители на хетерополизахаридите (гликозаминогликани) са хиалуронова киселина, хондроитинсулфат, кератинсулфат, дерматансулфат, хепарин. Гликозаминогликаните влизат в състава на протеогликаните (мукополизахариди), изграждат междуклетъчното пространство, задържат водата и обезпечават тургура на тъканите и еластичността на хрущялите. Изпълняват роля на смазочен и структурен компонент на ставите, хрущялите и кожата.
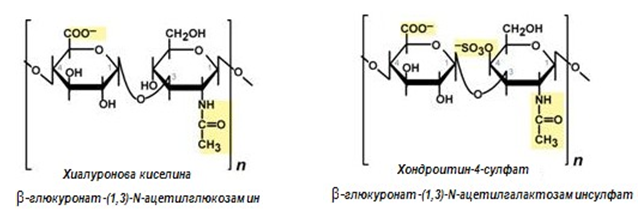
Хемицелулоза (полуцелулоза) - хетерополизахарид. В състава и влизат хексози, пентози и някои уронови киселини. Тя е доста застъпена в природата. Обикновено придружава същинската целулоза в стените на растителните клетки, но служи и като резервна храна.
Пектинови вещества - хетерополизахариди. Придружават целулозата в мембраната на месестите растителни части на цвеклото, морковите, и др., в зрелите плодове – ябълки, круши, лимони и др. Добива се чрез екстахиране с гореща вода на сок от плодовете. В присъствието на захари и органични киселини образува гели и затова намира приложение за приготвяне на плодови желета.
Хитин - принадлежи към скелетните полизахариди, които изграждат черупките на раци, външния скелет на членестоногите. Структурни елементи на хитина се явява N-ацетилглюкозамина, свързан помежду си с 1,4 – ß – гликозидни връзки, при пълна хидролиза се разпада до глюкозамин.
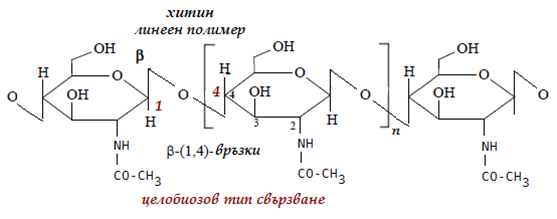
Фигура 74. Структура на хитин, целобиозов тип свързване, на N-ацетилглюкозамина, с β-1-4 гликозидни връзки
6.3. Въздействие на въглехидратите върху метаболизма
6.3.1. Гликемичен индекс
Различните видове въглехидрати въздействат по различен начин върху метаболизма. Това главно се дължи на скоростта, с която те се усвояват, измерваща се чрез така наречения гликемичен индекс и връзката му със секрецията на хормона инсулин. Гликемичният индекс показва колко се повишава нивото на кръвната захар при прием на определена храна. За еталон се приема глюкозата с гликемичен индекс 100 и останалите храни се определят спрямо нея. Например, белият хляб има гликемичен индекс около 70, което означава че приемът на определено количество бял хляб ще покачи нивото на кръвната захар на 70% спрямо същото количество глюкоза. Погрешно е разпространеното схващане, че простите въглехидрати имат висок гликемичен индекс, а сложните нисък. Богатите на нишесте картофи имат гликемичен индекс над 80, а фруктозата като представител на простите въглехидрати е с гликемичен индекс 22. Според гликемичния индекс въглехидратите се разделят условно в три групи:
- бавни – с индекс под 40;
- средни – между 40 и 60;
- бързи – индекс над 60.
Гликемичният индекс се отнася само за самостоятелната консумация на дадена храна. Различната комбинация от бавни, средни и бързи въглехидрати, значително го променя. Най-съществен принос за понижаване на гликемичният индекс, без да внасят допълнителни калории, имат фибрите. Те забавят скоростта на усвояване на сложните въглехидрати и могат да се добавят допълнително към храната или да се предпочитат храни, които ги съдържат в по-голямо количество. Поради значително повечето количество фибри в пълнозърнестия хляб, той е с гликемичен индекс 50, докато белият е с 70. Фрешовете също имат по-висок индекс от този на целите плодове, поради отсъствието на фибрите в първите.
| Продукт | Гликемичен индекс* |
| Глюкоза (еталон) | 100 |
| Мед | 87 |
| Картоф | 83 |
| Бял ориз | 82 |
| Корнфлейкс | 80 |
| Бял хляб | 70 |
| Заахар | 65 |
| Стафиди | 64 |
| Банан (зрял) | 62 |
| Царевица | 55 |
| Сладък картоф | 52 |
| Спагети | 50 |
| Пълнозърнест гляб | 50 |
| Пълнозърнести спагети | 40 |
| Портокал | 40 |
| Прясно мляко | 36 |
| Ябълка | 35 |
| Мляко | 35 |
| Боб | 33 |
| Грах | 33 |
| Леща | 29 |
| Грейпфрут | 25 |
Таблица 4. Гликемичен индекс на различни продукти
Всеки път, когато приемаме въглехидрати, нивото на кръвната захар се покачва. Това води до секрецията на хормона инсулин от панкреаса, който понижава нивото на кръвната захар, като усилва поемането й от клетките. Ако приемем въглехидрати с умерено висок гликемичен индекс, те ще се усвояват сравнително бавно и няма да доведат до рязко покачване на кръвната захар, следователно и секрецията на инсулин ще е умерена. Но при прием на бързи въглехидрати, те ще се усвоят много бързо и кръвната захар ще достигне сравнително високи стойности за кратко време. Това ще доведе до свръх отделяне инсулин, като реакция на организма да се справи веднага с хипергликемията. Вследствие на високите нива на инсулина, кръвната захар може да спадне дори под първоначалните си нива много скоро след приема на порцията бързи въглехидрати. Това ще ни подтикне към следващата порция бързи въглехидрати и така порочният кръг се завърта. Оттук произлиза и заключението, че сладкото отваря апетит за още сладко. Освен че увеличава концентрацията на глюкоза в мускулните и чернодробните клетки и способства за синтеза и натрупването на гликоген в тях, инсулинът стимулира и синтеза на мазнини, като вкарва излишните въглехидрати в мастните клетки. Също така инхибира ензима липаза, който е отговорен за освобождаването на мастните киселини от мастните депа. Не на последно място високите инсулинови нива потискат секрецията на растежния хормон, който освен всичко има и силно липотропно и мускулизграждащо действие.
Инсулинът е силно анаболен хормон – осигурява растежа като усилва поемането на аминокиселините от кръвта и използването им за синтеза на различни белтъци, особено в мускулите (болни от диабет деца остават ниски и дебели), също така потиска катаболния хормон кортизол.
6.3.2. Взаимопревръщане на въглехидратите в метаболизма
В тънките черва се всмукват всички видове монозахари постъпващи чрез храната (фруктоза, галактоза, маноза и т.н.), пред организма стои задачата, да превърнат получените хексози в глюкоза, за да може да бъде използвана в реакциите на метаболизма. Този процес е получил наименованието взаимопревръщане на захарите. Целта на този процес е да се получи само един субстрат α-D-глюкоза, което довежда до икономии на ресурси, тъй като се синтезират ензими за разграждане само за един субстрат, а не за всички видове монозахариди постъпващи в организма.
Превръщане на галактозата.
Галактозата отначало се превръща в галактозо-1-фосфат, от който се синтезира УДФ-галактоза, която изомеризира в УДФ-глюкоза. Източник на УМФ за синтезата на УДФ-галактоза се явява УДФ-глюкоза, която присъства в клетката (фиг.75).
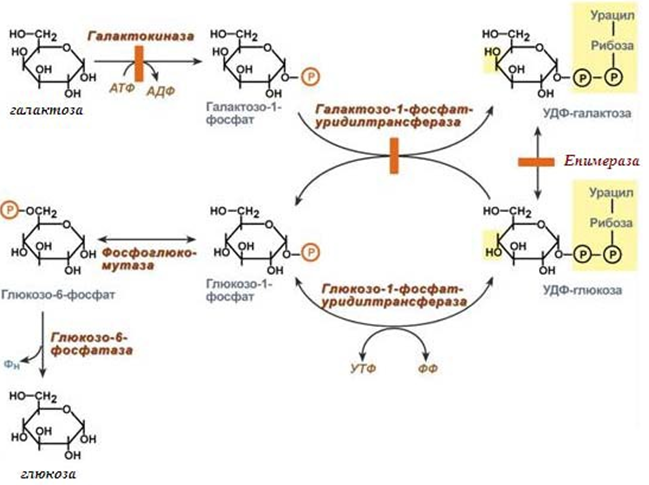
Фигура 75. Превръщане на галактозата в глюкоза
Синтезираната УДФ-глюкоза може да участва в:
- реакции на пренос на УМФ на галактозо-1-фосфат;
- да се превърне в свободна глюкоза и да влезе в кръвта;
- да се отправи за синтез на гликоген.
Превръщане на фруктозата.
Преобразуването на фруктозата до глюкоза се осъществява по две метаболитни вериги. В началото протича активация на фруктозата чрез фосфорилиране или на 6-ти въглероден атом, с участие на хексокиназата, или на 1-ви с участие на фруктокиназа (фиг.76).
Хексозокиназата има по-малко сродство към фруктозата и този път не е особено активен. Фруктозо-6-фосфата изомеризира в глюкозо-6-фосфат, следва отделяне на фосфатната група и образуване на глюкоза.
При образуване на фруктозо-1-фосфат, следва действие на алдолаза, която го превръща в глицералдехид и диоксиацетонфосфат, те се използват след това или в гликолизата, или в глюконеогенезата.
Особеност на метаболизма на фруктозата се явява, това че ензима фруктокиназа е инсулин-независим. По този начин, превръщането на фруктозата в пируват и ацетил-SКоА протича по-бързо, в сравнение с глюкозата. Игнорират се реакциите на регулиране и по-точно на лимитиране метаболизма на глюкозата, които се катализират от фосфоруктокиназата. Това води до по-високи нива на ацетил-SKoA, синтез на мастни киселини и натрупване на тлиацилглицероли.
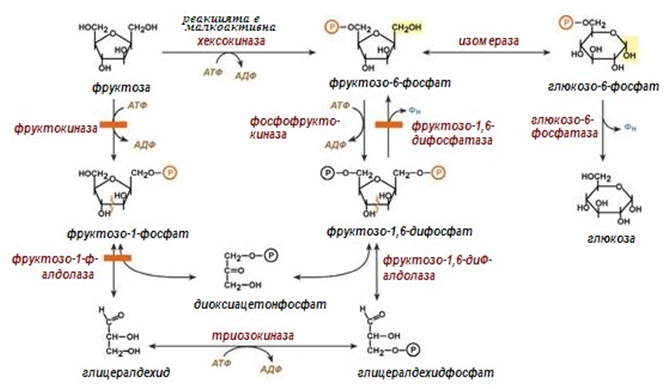
Фигура 76. Превръщане на фруктозата в глюкоза
Метаболизъм на глюкозата в клетката.
Глюкозата навлиза в клетката чрез кръвния поток. След постъпването в цитозола, глюкозата веднага се фосфорилира (фиг.77), под действие на ензима хексокиназа. След фосфорилирането глюкозата не може да напусне клетката, тъй като молекулата е отрицателно заредена, поради което ензима се нарича „капан на глюкозата“. Наличието на заредена група осигурява правилна ориентация на молекулата в активния център на ензима, освен това са намалява концентрацията на свободната нефосфорилирана глюкоза, което води до постъпване на нови молекули глюкоза от кръвта в клетката.
След фосфорилирането, т.нар. активиране на глюкозата, тя може да продължи по различни направления в метаболизма. Една част се използва за синтез на АТФ. При високи нива на глюкоза в клетките се синтезира гликоген, повечето тъкани са способни да синтезират гликоген. В хепатоцитите при високи кинцентрации и в адипоцитите, глюкозата се насочва към синтез на триацилглицероли, в черния дроб към синтез на холестерол. При определени условия част от глюкозата продължава по пентозофосфатния път, при който се образуват рибозо-5-фосфат и НАДФН. Определена част от глюкозата се използва за синтез на гликозамини и хетерополизахариди.
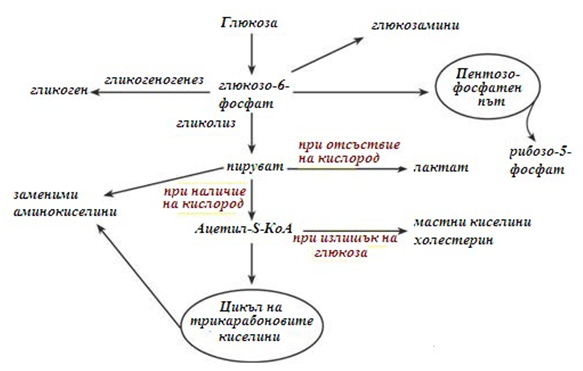
Фигура 77. Реакции на превръщане на глюкозата в клетката
Дефосфорилирането на глюкозата се осъществява от ензима глюкозо-6- фосфатаза. Този ензим съществува само в черния дроб и бъбреците. В клетките на черния дроб, наличието на ензима е свързано с регулационната функция на поддържане нивото на кръвната захар. В клетките на бъбреците, наличието на ензима е свързано с реабсорбцията на глюкозата. Двата вида клетки притежават способност да синтезират глюкоза (глюконеогенез).
Терминологичен речник
- #Въглехидрати – многоатомни алкохоли с алдехидна или кетонна група (полихидроксиалдехиди и полихидроксикетони), с обща емпирична формула (СН2О)6.
- #Алдози – алдехидната група е при краен С-атом, а при останалите С-атоми има по една –ОН-група. Според броя на въглеродните атоми биват: алдотетрози, алдопентози, алдохексози и т.н. По-голямо значение имат алдопентозите и алдохексозите, много от които са природни продукти.
- #Кетози – карбонилния С-атом (С=О) е най-често втория въглероден атом, според броя на въглеродните атоми се разделят на кетотетрози, кетопентози, кетохексози и т.н.
- #Дизахариди – изградени от по два монозахарида, свързването на две молекули монозахарид в молекула дизахарид става чрез обезводняване между две ОН-групи на монозахаридите, поне едната от които е гликозидна (гликозиден е този С-атом, който преди това е бил в състава на карбонилна група). Гликозидното свързване между монозахаридите, може да бъде трехалозов, малтозов, гентиобиозов и целобиозов тип свързване.
- #Гликозиден С-атом – който е бил в състава на карбонилна група.
- #Захароза - обикновена захар, изградена от една молекула глюкоза и една фруктоза, тя е α-D-глюкопиранозил-β-D-фруктофуранозид. Молекулата и е образувана чрез трехалозов тип 1-2 свързване на два гликозидни С-атома.
- #Малтоза - продукт от хидролизата на скорбялата и гликогена, които се разпадат по време на храносмилането, съставена е от две молекули глюкоза, свързани с α 1,4 – гликозидна връзка.
- #Лактоза - млечна захар, съдържаща се в млякото и някои млечни продукти. Изградена е от една молекула глюкоза и една галактоза.
- #Уронови киселини – карбоксилни производни, при които най-отдалечената от карбонилната група хидроксилна група е окислена до карбоксилова (уронови киселини).
- #Хетерополизахариди – характеризират се с повтарящи се дизахаридни остатъци, включват в състава си уронова киселина и аминозахар.
- #Полизахариди – изградени от стотици или хиляди монозахарида (мол. маса средно 100 000 Da), свързани чрез гликозидна връзка.
- #Аминозахари – една от хидроксилните групи (най-често тази на второ място) е заместена с аминогрупа.
- #Нишесте (скорбяла) – хомополимер на α-D-глюкозата. Молекулите на нишестето са изградени от α-D-глюкопиранозни остатъци, свързани чрез поликондензация от малтозов (1-4 свързване) и гентиобиозов тип (1-6 свързване). Нишестето е нееднородно и се състои от две вещества: амилоза (15-20%), и амилопектин (80-85%).
- #Амилоза. Молекулата на амилозата е изградена чрез 1-4 свързване на α-D- гликозидни остатъци, съдържа около 200– 1000 подобни остатъци.
- #Амилопектин. Молекулата на амилопектина е разклонена, изградена от α- D-гликозидни остатъци, свързани чрез 1-4 (малтозов) и 1-6 свързване (гентиобиозов тип).
- #Гликоген - резервен захарид, съставна част на нашия организъм, складова форма на глюкозата. Молекулната му структура е аналогична с тази на амилопектина, но е по-разклонена, средно на всеки 8-10 глюкозни остатъка има по едно 1-6 свързване и дължината на разклоненията са по-къси 11-18 глюкозни остатъци.
- #Целулоза – има линейна структура на молекулата, изградена от β-D- глюкопиранозни остатъци, свързани с β-1,4-гликозидна връзка, средният брой на глюкозидните остатъци в молекулата е 600-6000, а молекулната маса – от 100000 до 1000000.
- #Хитин - принадлежи към скелетните полизахариди, които изграждат черупките на раци, външния скелет на членестоногите. Структурни елементи на хитина се явява N-ацетилглюкозамина, свързан помежду си с 1,4 – ß – гликозидни връзки, при пълна хидролиза се разпада до глюкозамин.
- #Гликемичен индекс – показва с колко се повишава нивото на кръвната захар при прием на определена храна.

