

Ключови думи: нуклеозид, нуклеотид, полинуклеотидни вериги, ДНК, видове РНК (иРНК, рРНК, тРНК), нуклеопротеини, азотна база
Цели:
- Да се изясни съставът и структурата на нуклеиновите киселини.
- Да се разбират особеностите на полинуклеотидните вериги.
- Да се изяснят различията в строежа на ДНК и РНК.
- Да знаят особеностите на първичната структура на НК.
- Да се научат особеностите и конформацията на ДНК и на различните видове РНК (иРНК, рРНК, тРНК);.
- Да получат знания за биологичната роля на различните видове РНК.
Въведение
Биологичното значение на нуклеиновите киселини е съхраняване, реализиране и предаване на наследствената информация в потомството. До 40-те години на ХХ век за материален носител на наследствеността е считан белтъка, въпреки че дезоксирибонуклеинова киселина (ДНК) е открита във всички клетки от бактерии до човек, винаги съсредоточена в хромозомите.
Клетките съдържат два типа полинуклеотиди, ДНК и РНК. ДНК съхранява и предава наследствената информация, представлява двуверижна молекула изградена от две комплементарни полинуклеотидни вериги, служи като хранилище на генетична информация. Двуверижният строеж осигурява по- висока надеждност на съхраняваната информация. При репаративен процес неповредената ДНК молекула се използва като матрица за коригиране и възстановяване на повредената комплементарна верига.
Молекулите на РНК, които са едноверижни, изпълняват други функции:
- участват в синтезата на специфични белтъци;
- изпълняват структурна функция.
Нуклеиновите киселини свързани с белтъци (нуклеопротеини), изграждат рибозоми, хроматин, вируси. В рибозомата рибонуклеиновата киселина (РНК) е свързана със специфични (рибозомни) белтъци. Вирусите са практически чисти рибо- или дезоксирибонуклеопротеини. Хроматина е изграден от хистонови и нехистонови белтъци и дезоксирибонуклеинова киселина.
8.1. Наследственост и поток на генетичната информация – централна догма на молекулярната биология
Нуклеиновите киселини са биополимери открити през 1868г. от швейцарския биохимик Фридрих Фишер в ядрата на гнойни клетки. Поради киселинния си характер са наречени нуклеинови (лат. нуклеус – ядро) киселини. До 40-те години на ХХ век господства схващането, че материалният носител на наследствеността е белтъкът, въпреки че ДНК (дезоксирибонуклеинова киселина) е открита във всички клетки растителни и животински, във всички организми от бактериите до човека, винаги съсредоточени в ядрото и в хромозомите.
Доказването на генетичната роля на ДНК, се предхожда от някои открития:
- 1928г. Грифит (англ. бактериолог) открива трансформацията при пневмококите;
- 1944г. О.Ейвъри, Маклеод и Маккарти, доказват че материалния носител на наследствените белези при пневмококите е ДНК;
- 1952г. Хърши и Чейз доказват при бактериофаги, че носител на наследствените свойства е само ДНК, но не и белтъци;
- 1956г. Френкел-Конрат с ВТМ доказва, че при растителния вирус носител на наследствеността е РНК.
През 50-те години на ХХ век е доказано, че материалния носител на наследствената информация са нуклеиновите киселини. Биологичното им значение е запазване и предаване на наследствените качества в поколенията (ДНК) и реализирането им в индивидуалния живот (ДНК и РНК).
Нуклеиновите киселини са основен компонент на живата клетка. Според вида на клетките (доклетъчните форми), в чието изграждане участват, биват:
- Вирусни нуклеинови киселини (1- или 2- верижна РНК или 1- или 2- верижна ДНК);
- Бактериални ДНК (нагъната или кръгова форма) и РНК;
- Еукариотни НК;
При вирусите се среща само по един вид нуклеинова киселина (ДНК или РНК). В еукариотните клетки по-голямата част от генетичния материал е съсредоточена в клетъчното ядро - над 80% от ДНК, и в него се синтезират всички РНК (иРНК, рРНК и транспортна - тРНК). Освен в ядрото ДНК се съдържа и в митохондриите и пластидите - органелите, които снабдяват клетката с енергия и се делят независимо от клетката.
Нуклеиновите киселини са хетеробиополимери с изключителна биологична роля, отразена в т.н. централна догма на молекулната биология, формулирана от Франсис Крик . "Централна догма“ от Ф. Крик отразява ролята на нуклеиновите киселини за съхраняване и предаване на наследствената информация.
Потока на генетичната информация е строго детерминиран и се изразява чрез централната догма на молекулярната биология, съгласно която преноса на генетична информация се осъществява единствено от нуклеиновите киселини (ДНК и РНК). Получател на генетичната информация може да бъде друга нуклеинова киселина (ДНК или РНК) и белтък.
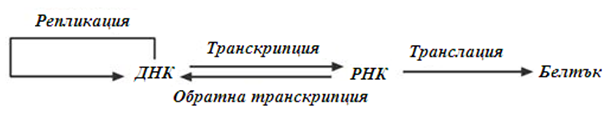
Фигура 102. Централна догма на молекулярната биология
Предаването на наследствената информация, в се осъществява от майчината клетка към дъщерната, с помощта на ДНК (репликация). За използване на генетичната информация е необходима РНК, която се образува от матрицата на ДНК (транскрипция). РНК непосредствено участва във всички етапи на синтеза на белтъчни молекули (транслация), което обезпечава структурата и функционирането на клетката. В някои вируси е възможен и обратен поток на информацията от РНК към ДНК, т.нар. процес обратна транскрипция.
8.2. Пурини и пиримидини. Нуклеозиди, нуклеотиди и полинуклеотиди
Нуклеиновите киселини са полимери (хетеробиополимери), изградени от мономери, наречени нуклеотиди, които са по-сложни от аминокиселините – мономери на белтъците. Всеки нуклеотид съдържа фосфорна киселина (един, два или три остатъка), монозахарид (пентоза – рибоза или дезоксирибоза), азотна база от пуриновия ред (аденин, гуанин) или от пиримидиновия (цитозин, урацил или тимин).
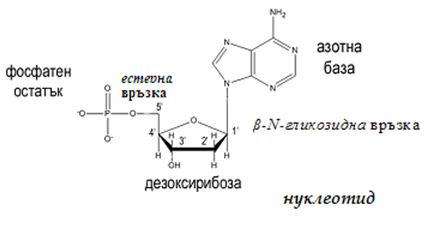
Фигура 103. Строеж на нуклеотид
Азотната база се свързва с пентозния остатък чрез β-N-гликозидна връзка, като пиримидиновата база се свързва при N-1, а пиридиновата при N-9. N-гликозидната връзка в нуклеозидите се получава при обезводняване на цикличната NH-група при N9 в пуриновата база (или N1 в пиримидиновата база) и гликозидната НО-група при С1'-атом в пентозата. Остатъците на фосфорната киселина се свързват естерно на 3’- и/или 5’- позиция с пентозния остатък. В молекулите на нуклеиновите киселини захарният остатък винаги е с петатомен пръстен (пентоза). В зависимост от това какъв монозахариден остатък участва в изграждането на полинуклеотидната верига има два типа нуклеинови киселини: дезоксирибонуклеинови (ДНК) и рибонуклеинови (РНК). ДНК съдържат дезоксирибоза, а РНК – рибоза (фиг.104).
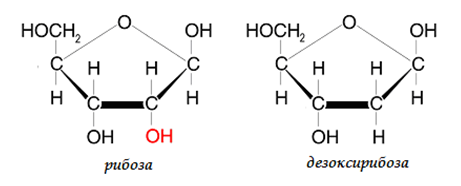
Фигура 104. В молекулите на нуклеиновите киселини захарният остатък винаги пентоза, ДНК съдържат дезоксирибоза, а РНК - рибоза.
ДНК е доста стабилна в химично отношение, защото в пентозата й при 2’ въглероден атом липсва –ОН група, а и базите й са с хидрофобен характер. Монозахаридният остатък (рибоза или дезоксирибоза) се свързва с азотна база. Азотните бази са пръстеновидни молекули, които съдържат освен въглеродни атоми (като органични съединения) още и азотни атоми. Те биват две пуринови (големи) бази – аденин (А) и гуанин (Г) и три пиримидинови (малки) бази – тимин (Т) (урацил при РНК) и цитозин (Ц) (фиг.105).
Номерацията на атомите в 6-атомния пуринов пръстен е обратно на часовниковата стрелка , а в 5-атомния пръстен по часовниковата стрелка. Номерацията в пиримидиновия пръстен е по часовниковата стрелка.
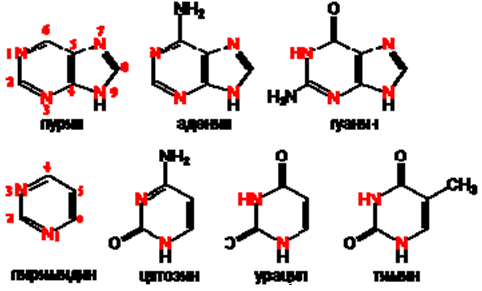
Фигура 105. Структура на главните азотни бази в нуклеиновите киселини: две пуринови - аденин (А) и гуанин (Г)
три пиримидинови - цитозин (Ц), урацил (У) и тимин (Т).
При свързване на базата с пентоза чрез N-гликозидна връзка се образува нуклеозид.
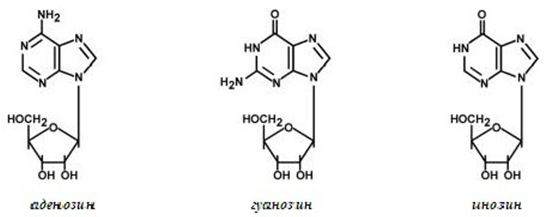
Фигура 106. Структура на пуриновите нуклеозиди
Пуриновите нуклеозиди имат окончание -озин (аденозин, гуанозин, фиг.106), а пиримидиновите имат окончание -идин (уридин, тимидин, цитидин, фиг.107).
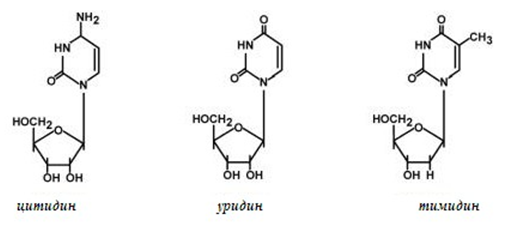
Фигура 107. Структура на пиримидиновите нуклеозиди
При свързване на нуклеозид с фосфорна киселина се получава нуклеотид. Връзката между пентозата на нуклеозида и фосфорната киселина е естерна (фиг.103).
Нуклеотидите са градивните единици на нуклеиновите киселини. Освен това като свободни нуклеотиди имат съществени функции:
- Нуклеозид-дифосфатите и -трифосфатите съдържат киселинно- анхидридни фосфатни връзки, които са макроергични или богати на енергия връзки, с които осигуряват енергия на биосинтезните и други ендергонични процеси.;
- Участват в метаболизма;
- Влизат в състава на по-сложни коензими: никотинамидаденин динуклеотид (НАД), никотинамидаденин динуклеотидфосфат (НАДФ), флавинадениндинуклеотид (ФАД) и коензим А (КоА).;
- Имат регулаторен ефект, напр. съотношението АДФ/АТФ регулира скоростта на окисление в митохондрийната дихателна верига. Някои нуклеотиди действат като алостерични ефектори (инхибитори или активатори) върху различни ензимни активности.;
- Цикличните нуклеотиди цАМФ и цГМФ участват в трансдукцията или препредаването на хормонални сигнали, а също и като активатори на транскрипция на някои гени.
При свързване на нуклеотидите се получава т.нар. “захарофосфатен скелет”. Монозахаридният остатък взаимодейства с фосфорната киселина и формира захарофосфатен скелет на нуклеиновата киселина. Нуклеотидите са свързани чрез 3',5'-фосфодиестерни връзки. Захарофосфатният скелет в ДНК и РНК е отрицателно зареден, поради наличието на фосфатни групи. Наличието на фосфатен остатък придава киселинните свойства на нуклеиновите киселини.
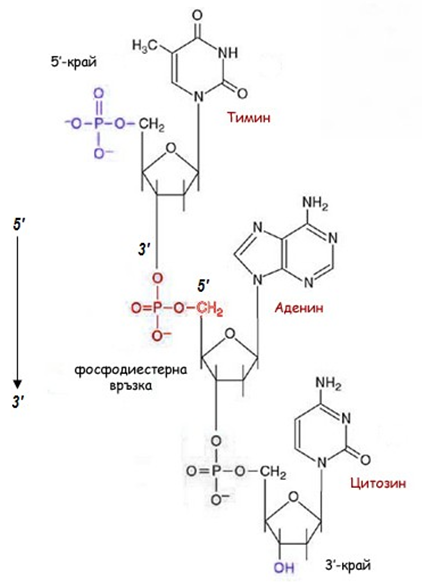
Фигура 108. Първична структура на полинуклеотидна ДНК верига (фосфорната киселина при 5’С-атом естерифицира с -ОН от 3’С-атом), образува се 3',5'-фосфодиестерна връзка.
НК са разнообразни по строеж и големина съединения. В ДНК участват А, Г, Ц и Т. В РНК участват А, Г, Ц и У. Разнообразието им се определя от подреждането на азотните бази на отделните нуклеотиди в състава на полинуклеотидната верига по точно определен ред, т.е. от първичната им структура. Това от своя страна определя индивидуалността на всяка конкретна молекула НК. Първичната структура определя биологичните свойства на нуклеиновите киселини.
Всяка от веригите е изградена от свързани помежду си мономерни единици (нуклеотиди) чрез ковалентни фосфодиестерни връзки. Фосфорната киселина на единия нуклеотид при 5’ въглероден атом естерифицира хидроксилната група от 3’ въглероден атом на пентозата на следващия нуклеотид. Веригата на нуклеиновата киселина е отворена (при еукариотите), и винаги в единия край – 5’ е с фосфатна група, а в другия край – 3’ е със свободна –ОН група (фиг.108).
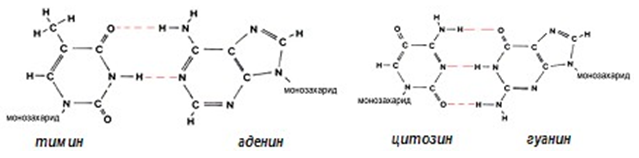
Фигура 109. Комплементарност между азотните бази,свързани чрез водородни връзки
Полинуклеотидните вериги на ДНК и РНК притежават направление, т.е. имат 3’ и 5’ край. В ДНК веригите са антипаралелни, т.е. направленията са в обратни посоки. Съществува и комплементарност при азотните бази, което се обуславя от тяхната структура, съответно аденин е комплементарен с тимин (А=Т), а гуанин е комплементарен с цитозин (Г=Ц).
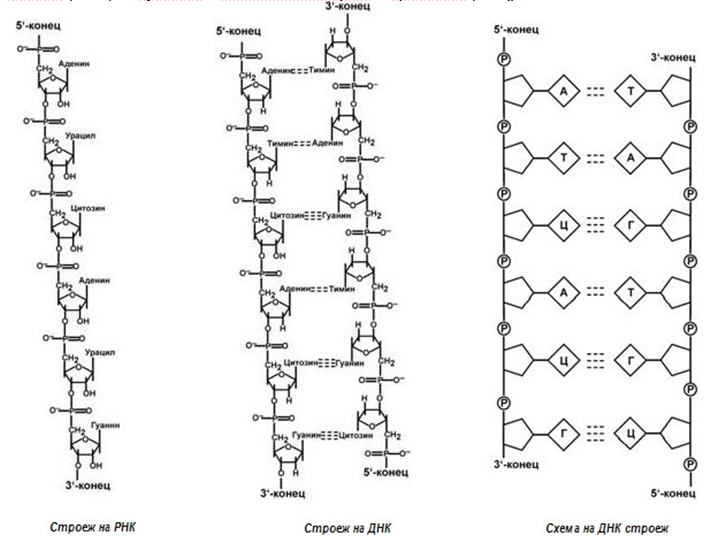
Фигура 110. Различие в структурите на РНК и ДНК
Различия между ДНК и РНК (фиг.110):
- количество вериги – при РНК е една, при ДНК са две;
- размери – ДНК е по-голяма молекула;
- локълизация в клетката – ДНК се намира в ядрото, РНК почети цялото количество е извън ядрото;
- монозахарид – в ДНК – дезоксирибоза, в РНК – рибоза;
- азотна база – в ДНК има тимин, в РНК – урацил;
- функция – ДНК съхранява генетичната информация, РНК я реализира.
8.3. ДНК - носител на генетичната информация.
Основните биологични функции на ДНК са да съхранява и предава генетичната информация за клетъчната програма. Дезоксирибонуклеиновата киселина е молекула на наследствеността. Установено е, че молекулата на ДНК представлява спирална структура, изградена от две полинуклеотидни вериги. Именно двойната спирала на ДНК е структурата, в която е записана цялата наследствена информация (програма) на всяка жива клетка.
Едно вещество може да бъде материален субстрат на наследствеността, ако едновременно отговаря на следните условия:
- Строежът на молекулите му да позволява записване и запазване на наследствената информация.
- Количеството му в клетките да е постоянно, да не се променя или да не изчезва и да не се синтезира наново.
- Да се самовъзпроизвежда съвсем точно и новите копия да се разпределят по равно във всички клетки и във всички индивиди. Това осигурява предаването на наследствената информация.
Индивидуалността на нуклеиновите киселини се определя от подреждането на четирите нуклеотида по дължината на полинуклеотидната верига. Наследствената информация е информация за цялостната биологична характеристика на клетката, за нейните структурни и функционални особености. Тя е записана под формата на трибуквени означения – кодони. Кодонът представлява определена последователност от три нуклеотида от веригата на ДНК. Участъкът от ДНК, който носи информация за синтезата на белтък (или за синтезата на една иРНК) се нарича ген.
Редица изследвания показват, че гените са участъци от молекулата на ДНК и контролират синтезирането на специфични белтъци. В един ген е програмирано синтезирането на една полипептидна верига. Генът е най- малката функционална единица за наследственост. Днес се използват няколко определения за ген:
- последователност от ДНК, отговорна за синтезата на една полипептидна верига или на една иРНК;
- последователност от ДНК, която се презаписва.
Ето защо наследствената програма се нарича още генетична програма или генетична информация. Начинът на записване на наследствената информация в молекулата на ДНК се нарича генетичен код.
Дезоксирибонуклеиновите киселини (ДНК) съхраняват и предават наследствената информация от едно поколение на друго, а освен това определят свойствата на живата клетка чрез регулиране експресията на генетичната информация, главно чрез упражняване на контрол върху синтезата на рибонуклеинови киселини (РНК) и белтъци.
8.4. Двойна спирала. Принцип на комплементарност.
Нуклеиновите киселини са разнообразни по строеж и големина съединения. Разнообразието им се определя от подреждането на азотните бази на отделните нуклеотиди в състава на полинуклеотидната верига по точно определен ред. Това от своя страна определя индивидуалността на всяка конкретна молекула ДНК.
Азотните бази са насочени навътре от захарофосфатния скелет на молекулата и са като че ли скрити в нея. Те са свързани помежду си чрез
водородни връзки (фиг.110). Така подредените бази могат и допълнително да взаимодействат помежду си и да допринесат за стабилността на молекулата. Нуклеиновите киселини са стабилни, но и изменчиви молекули, както в структурно, така и в химично отношение. Само съчетанието на стабилност с известна изменчивост може да осигури изпълнението на сложните биологични функции на нуклеиновите киселини. Полинуклеотидната верига има посока – единият край има свободна – ОН група на рибозата, а другият – фосфорна киселина.
Двете полинуклеотидни вериги, формиращи спиралата, образуват при спирализирането си две бразди – малка и голяма (фиг.111).
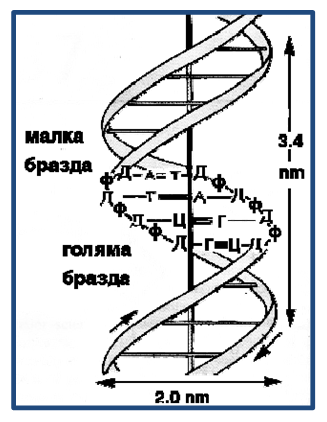
Фигура 111. Модел на Уотсън и Крик за двойно-спиралната структура на ДНК.Ширината на дясно-въртящата двойна спирала е 2.0 nm;Ходът на спиралата е 3.4 nm; В един ход на спиралата има 10.5 нуклеотидни двойки; Дебелата вертикална линия представлява централната ос на спиралата; Веригите са антипаралелни и комплементарни.
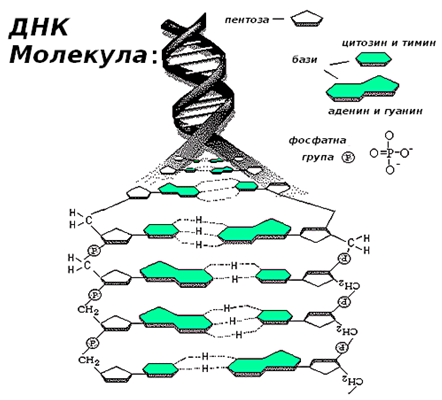
Фигура 112. Структура на ДНК – двойна спирала
Вторичната структура на ДНК – двойна спирала (фиг.112), е открита през 1953г. от Уотсън и Крик, благодарение на откритото правило на Чаргаф за комплементарност (допълнителност) на базите: сумата А+Г = Ц+Т, А=Т и Ц=Г. Основна отличителна черта на двойната спирала е, че при нейното изграждане една пуринова база винаги се свързва с една пиримидинова база от срещуположната полинуклеотидна верига:
- базата аденин се свързва чрез две водородни връзки с базата тимин;
- гуанин се свързва чрез три водородни връзки с базата цитозин.
Така се спазва точно определената съответност (комплементарност). Тази комплементарност предопределя начина, по който се синтезира (удвоява) молекулата ДНК.
Благодарение на строежа и на комплементарността (допълнителността) на двете съставящи ДНК вериги е осигурено условието - молекулата може по съвсем точен начин да се самовъзпроизвежда (да се реплицира). Процесът се нарича репликация на ДНК.
8.5. Репликация - удвояване на генетичната информация.
През 1953г. е направено едно от най-вълнуващите открития в биологията – откритието на Уотсън и Крик за структурата на двойната спирала на ДНК. От свързването на азотните бази непосредствено следва и механизмът, по който може да се копира записаната в ДНК генетична информация. При разделянето си на две полинуклеотидни вериги, всяка една става модел (матрица), по която откопира огледалният си образ.
Репликацията (синтез) на ДНК протича в строго определен период от живота на клетката. Клетъчния цикъл има 4 фази (фиг.113):
- митоз (М);
- синтетична фаза (S);
- предсинтетична (G1 – от англ. gap - интервал);
- постсинтетична (G2).
Важно участие в регулиране смяната на фазите имат циклините – белтъци с маса 90кДа, които активират субединиците на ензими известни като циклин-зависими кинази (ЦЗК). Активните комплекси на циклин-ЦЗК фосфорилират вътреклетъчните белтъци като изменят тяхната активност. По такъв начи обепечават смяната на фазите в клетъчния цикъл.
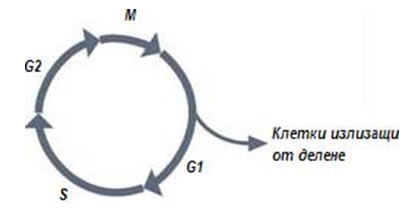
Фигура 113. Фази на клетъчения цикъл - митоз (М), синтетична фаза (S), предсинтетична (G1 – от англ. gap - интервал) и постсинтетична (G2)
Механизмът на репликация е установен от Матью Мезелсон и Франклин Стал през 1957г. (фиг.114). Синтезът (репликация, удвояване) на ДНК протича през S–фазата на клетъчния цикъл, когато клетката се подготвя за делене. Механизмът на репликация е полуконсервативен, т.е. всяка материнска верига синтезира дъщерно копие.
Репликацията се осъществява като двойната спирала на молекулата се разплита под действието на специални ензими, които разкъсват водородните връзки между срещулежащите бази. Върху всяка единична верига започва да се синтезира допълващата я (комплементарна) верига. При това се спазват правила за комплементарност (допълнителност): срещу аденина винаги застава тимин, а срещу тимина — аденин, срещу гуанина — цитозин, и срещу цитозина — гуанин. Всяка единична верига от ДНК служи като своеобразна матрица, която определя последователността на нуклеотидите в новосинтезираната верига.
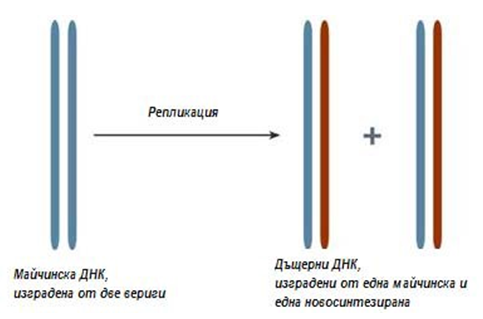
Фигура 114. Полуконсервативен механизъм на репликация
При делене на клетката, молекулата на ДНК се удвоява, като ДНК се деспирализира от определено място, наречено ori (от англ. origin – начало). Репликацията на ДНК се осъществява въз основа на матричния принцип и по принципа на комплементарността,т.е. срещу всеки нуклеотид, съдържащ определена азотна база, застава съответният му комплементарен нуклеотид. Термина матричен биосинтез или матричен принцип, означава че клетката синтезира полимерни молекули, такива като нуклеинови киселини и белтъци, на основата на шаблон – матрица. Това обезпечава точното предаване на сложните структури на съществуващите молекули в новосинтезираните. Така последователността на нуклеотидите в ДНК диктува подреждането на нуклеотидите в новосинтезиращата се верига. Отделните нуклеотиди се свързват и образуват полинуклеотидна верига. Всяка новоизградена верига ДНК копира структурата на верига от старата молекула.
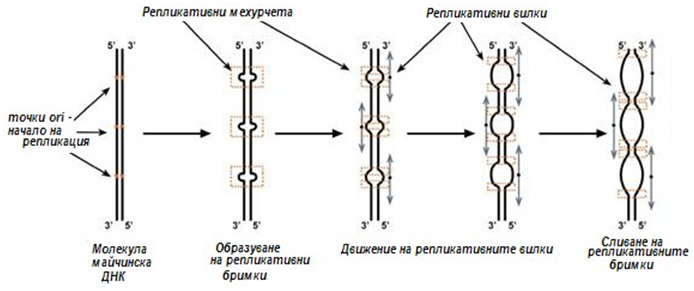
Фигура 115. Схема на възникване на репликативни „мехурчета“ (бримки), стрелките показват посоката на движение на репликативните вилки
Репликацията на ДНК е сравнително бърз процес. При бактериите тя започва от една точка и протича последователно (пълзи) по цялата дължина на молекулата. Скоростта е 30 mm/min, което означава, че за 1 s се изгражда нов участък от 500 нуклеотида. Еукариотните организми имат много по-дълги молекули ДНК в сравнение с прокариотните. При еукариотите репликацията се извършва по същия механизъм, но започва едновременно от няколко точки (до 100 при млекопитаещите). Молекула ДНК при еукариотите е съставена от много едновременно реплициращи се участъци, наречени репликони.
Репликацията протича в двете посоки от началната точка, оформя се репликативно „мехурче“, в което с формират две репликативни „вилки“, в тях протича разплитане, развъртане, синтез на ДНК. При еукариотните репликацията завършва за 9 часа.
Синтезът на новата ДНК верига протича в направление от 5’ към 3’ края, т.е. 5’-края на новата ДНК остава свободен, следващия нуклеотид се присъединява към 3’-ОН група на предходния нуклеотид, със скорост около 100 броя в секунда (за еукариотите). В репликативната“вилица“, в посока 5’→3’, процеса е непрекъснат, синтезира се нова верига ДНК наречена водеща. Направлението 5’→3’ е противоположно на посоката на движение на репликативната вилица, тук синтезата протича прекъснато на отделни фрагменти наречени фрагменти на Оказаки, а веригата която се синтезира се нарича изоставяща.
Репликацията на ДНК, протича по реда:
- ДНК-топоизомерази, разполагат се пред репликативната вилица, разрязват молекулата на ДНК, като облегчават разплитането и развиването;
- ДНК-хеликази, следват топозомеразите, развиват и разплитат молекулата ДНК;
- ДНК-свързващи белтъци, свързват разплетените вериги ДНК и ги стабилизират, като не допускат обратното им „слепване“ една с друга;
- ДНК-полимераза δ (гр. δ – делта), осъществява синтез на водещата верига дъщерна ДНК в направление 5’→3’ на матрицата на майчината ДНК верига по направление от нейния 3’-край към 5’-край (със скорост 100 двойки нуклеотиди в секунда).
Репликацията на изоставящата верига протича по-особено:
- Непосредствено след разплитането и стабилизацията на веригите на майчината молекула ДНК, към другата верига се присъединява ДНК- полимераза α и в направление 5’→3’ се синтезира праймер (РНК- праймер) с дължина от 10 до 200 нуклеотида. След което ензима се отделя от ДНК нишката.;
- Вместо ДНК-полимераза α към 3’-края на праймера се присъединява ДНК-полимераза ε (гр.ε-епсилон), продължава синтезата на дъщерната верига, но с дезоксирибонуклеотиди (с дължина до 150-200 нуклеотида). В резултат се образува нишка от две части РНК (т.е.праймер) и ДНК. ДНК-полимераза ε синтезира ДНК верига, докато не стигне до РНК-праймер, на предходния фрагмент на Оказаки (синтезиран малко преди това). След което ензима се отделя от веригата.
- ДНК-полимераза β (гр.β–бета), застава на мястото на ДНК- полимераза ε, предвижва се в направление 5’→3’ и отделя РНК- праймера, едновременно с това вгражда дезоксирибонуклеотиди на тяхно място. Ензима работи до пълното отделяне на праймера, докато не достигне ДНК, която е синтезирана по-рано от ДНК-полимераза ε. Ензима не може да се свърже с този сегмент и напуска нишката. В резултат върху матрицата майчина верига „лежи“ фрагмент дъщерна ДНК, която се нарича фрагмент на Оказаки.
- ДНК-лигаза, сшива два съседни фрагмента на Оказаки, т.е. 5’-края на участъка синтезиран от ДНК-полимераза ε, и 3’-края на веригата изграден от ДНК-полимераза β.
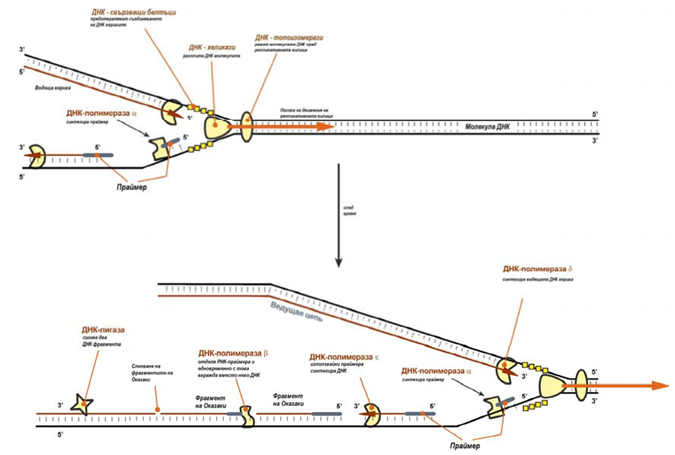
Фигура 116. Схема на репликативна „вилица“ – роля на ензимите
При просто устроените клетки ДНК е гола молекула. С усложняването на клетъчната структура, тя вече е “облечена”, свързана с белтъци. Колкото повече се усложнява клетката, толкова повече информация трябва да се съдържа в нея. Молекулата на ДНК нараства и трябва да е компактна, за да се побере в клетъчното ядро.
8.6. PHK – структура и видове. Транскрипция.
Мономери за изграждане на рибонуклеиновите киселини са монозахарид рибоза, а вместо азотната база тимин (Т) участва базата урацил (У). В РНК има свободна –ОН група на рибозата. Това води до по-голяма химическа нестабилност на молекулата в сравнение с тази на ДНК. Във воден разтвор РНК се разкъсва много по-бързо, отколкото ДНК. Ето защо ДНК е по- подходяща за запазване на наследствената информация от РНК.
Полинуклеотидната верига на РНК е едноверижна. Основните градивни частици на РНК са рибонуклеотидите, съставени от остатък на фосфорна киселина, азотна база и рибоза.
Известни са три вида РНК:
- Информационна;
- Транспортна;
- Рибозомна.
Веригите на рибонуклеиновите киселини най-често са едноверижни като само отделни участъци от единствената полинуклеотидна верига, огъвайки се могат да образуват двойноспирални участъци. Образуването на двойноспиралните участъци става между комплементарните бази – пиримидинова с пуринова (А с У и Г с Ц), както при ДНК.
В най-голямо количество са представени рибозомните РНК (рРНК) – 80% от клетъчната РНК. Тези РНК изграждат специализираните клетъчни органели – рибозомите, където се извършва синтезата на белтъците.
Информационните РНК (иРНК) представляват около 2% от РНК. иРНК презаписва информацията, носена от ДНК. В този процес се откопира последователността на базите от ДНК и презаписаната информация се пренася до рибозомите. Там тази информация се разчита и по нея се синтезират клетъчните белтъци. Информационните или матрични РНК (иРНК) пренасят информацията за синтеза на белтъци от ДНК в ядрото към специални органели в цитоплазмата, наречени рибозоми, където се извършва белтъчната биосинтеза.
Третият тип РНК е транспортната РНК (тРНК). Представлява около 15% от клетъчната РНК. Тази РНК има много специфична структура, приличаща на детелинов лист (фиг.117).
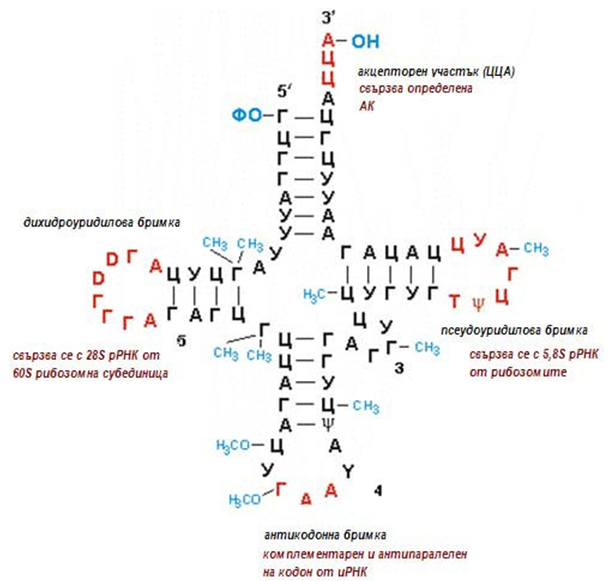
Фигура 117. Схема на структура на тРНК
Молекулата й е така устроена, че от една страна могат да се свързват с определени аминокиселини, а от друга страна да се свързват и с иРНК. Названието “транспортна” идва от функцията, която тези РНК изпълняват – пренос на аминокиселините до рибозомите, където се намира иРНК. Това са молекули-посредници, които осъществяват връзката между презаписаната информация в иРНК и подреждането на аминокиселините – мономерната градивна единица на белтъчните молекули. С други думи, с тяхна помощ се осъществява превеждането на генетичния запис на езика на белтъците.
Във всяка тРНК има четири важни участъка (фиг.117):
- акцепторен участък (тройка нуклеотиди ЦЦА в 3'-края), където се свързва определена аминокиселина;
- псевдоуридилова бримка (Т-ψ-Ц) - за свързване с 5,8S рРНК от рибозомите;
- антикодонна бримка, съдържа специфичен триплет нуклеотиди, наречен антикодон, тъй като е комплементарен и антипаралелен на определен кодон в иРНК;
- дихидроуридилова бримка (D). Тя се свързва с 28S рРНК от 60S рибозомната субединица
Всяка тРНК носи определена аминокиселина, когато се разчете последователно информационния запис, аминокиселините се свързват помежду си и образуват полипептидна верига (транслация).
Преди да се синтезират белтъци, информацията за техния строеж трябва да се вземе от ДНК и да се достави до мястото на синтеза на белтъците. С това са занимават информационните или матрични РНК. Едновременно с това на клетката са необходими транспортни РНК и също така структурните компоненти на рибозомите, които синтезират белтък – рибозомни РНК. Пълната информация за строежа на транспортните и рибозомните РНК също се намира в ДНК. Процесът на презаписване на данните от ДНК на РНК е транскрипция (англ. transcription – презаписване) – биосинтез на РНК от матрицата на ДНК.
За протичане на транскрипцията са необхидими:
- матрица – една от нишките ДНК;
- растяща верига – РНК;
- субстрат за синтез – рибонуклеиди (УТФ, ГТФ, ЦТФ, АТФ);
- източник на енергия – УТФ, ГТФ, ЦТФ, АТФ;
- ензими РНК-полимерази и белтъчни фактори за транскрипция.
Биосинтезата на РНК протича в участък на ДНК, който се обозначава като транскриптон, от единия край той е ограничен от промотора (начало), а от другия с терминатора (края).
Транскрипцията протича в три етапа (фиг.118).:
Инициация – промотора съдържа стартов сигнал за транскрипция – ТАТА-бокс. Така се нарича определена последователност от нуклеотиди в ДНК, свързващи първия фактор на инициацията ТАТА – фактор. Този фактор обезпечава присъединяването на РНК-полимераза към ДНК нишката, която ще се използва в качество на шаблон за транскрипиция (матричната нишка ДНК). Промотора е асиметричен и свързва РНК-полимеразата само в една ориентация, така се определя посоката на транскрипция, в направление от 5’-края към 3’-края (5’→3’). За свързването на РНК-полимеразата с промотора е необходим още един фактор за инициация σ-фактор (гр. σ-„сигма“), веднага след формирането на РНК промотора (с дължина 8-10 рибонуклеотида), σ-фактора се отделя от ензима. Други фактори развиват ДНК веригата пред РНК-полимеразата.
Елонгация – белтъчните фактори за елонгация обезпечават предвижването на РНК-полимеразата по дължина на ДНК нишката и разплитат молекулата ДНК по дължина до 17 нуклеотидни двойки. РНК- полимеразата се предвижва със скорост 40-50 нуклеотида в секунда в направление 5’→3’. Ензимът използва АТФ, ГТФ, ЦТФ, УТФ едновременно в качеството на субстрати и в качество на източник на енергия.
Терминация – РНК-полимеразата, когато достигне терминиращия кодон спира. С помощта на белтъчния фактор за терминация, така наречения ρ- фактор (гр. ρ-„ро“), от матрицата на ДНК се отделят ензима и синтезираната молекула РНК, която е така наречения първичен транскриптон, предшественик на иРНК или тРНК, или рРНК.
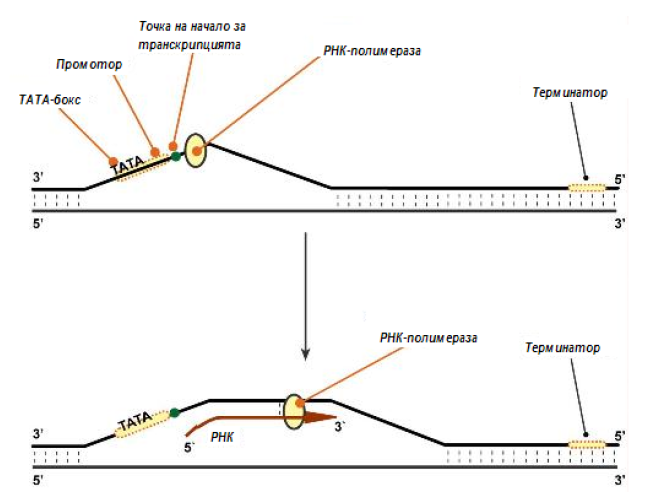
Фигура 118. Схема на процеса транскрипция
Новосинтезираните РНК са неактивни („незрели“), претърпяват редица изменения, които са известни като процесинг. При еукариотите на процесинг са подложени всички видове пре-РНК, докато при прокариотите единствено предшествениците на рРНК и тРНК.
При транскрипцията на иРНК, се образуват хетерогенни ядрени РНК, които по размер са по-големи от иРНК. Тези хетерогенни РНК включват информативни (екзони) и неинформативни (интрони) участъци. Протича процес известен като сплайсинг (англ.splice – снаждам въже), при който с участие на малки ядрени РНК се отделят интроните и се свързват екзоните.
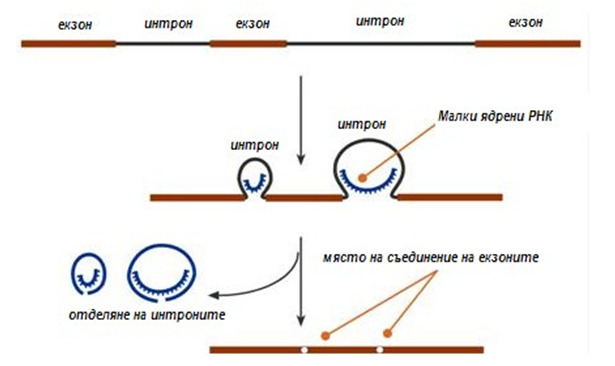
Фигура 119. Схема на процеса сплайсинг
По време на транскрипцията към пре-ИРНК се присъединява на т.нар. “шапка“, коята защитава молекулата РНК от екзонуклеазите работещи с 5’- края и също така свързва иРНК с рибозомата при началото на транслацията. Към 5’-края на пре-иРНК се прикрепва N7-метил-гуанозин. С помощта на полиаденилат-полимераза и АТФ, протича присъединяване на 100 до 200 аденилови нуклеотиди, към 3’-края на РНК, т.нар. поли(А)-опашка. Поли(А)- опашката е необходима за защита на молекулата РНК от екзонуклеазите, работещи с 3’-края.
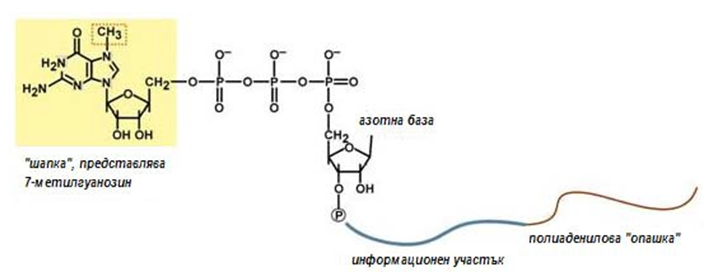
Фигура 120. Схема на зряла молекула иРНК след процесинг
Предшествениците на рРНК са по-големи молекули в сравнение със зрелите рРНК. При тяхното зреене прерибозомната РНК се разрязва на по- малки форми, които непосредствено участват във формирането на рибозомите. При еукариотите съществуват четири типа рРНК – 5S-, 5,8S-, 18S- и 28S-рРНК. При това 5S-рРНК се синтезира отделно, а голямата прерибозомна 45S-рРНК се разпада на 5,8S-, 18S- и 28S-рРНК. При прокариотите молекулите рибозомни РНК имат различни свойства (5S-, 16S- и 23S-рРНК).
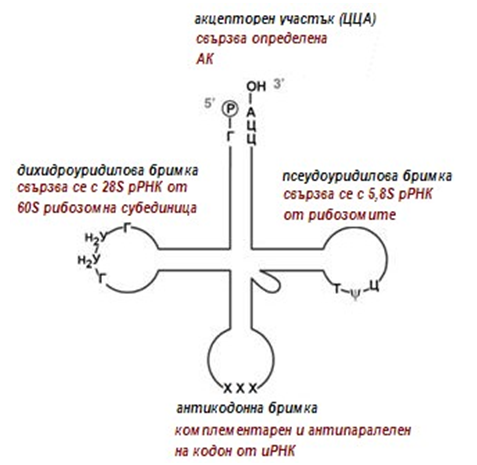
Фигура 121. Схема на вторичната структура на зряла молекула тРНК след процесинг
Зреенето на предшественика на тРНК се изразява в модификация на нуклеотидите в молекулата по пътя на дезаминиране и метилиране, като се образува псевдоуридин и дихидроуридин. Формира се антикодоновата бримка като по пътя на сплайсинга се отделя интрона от средната част на пре-тРНК. Формира се в 3’-края последователността ЦЦА. При това при някои пре-тРНК от 3’-края се отделят излишните нуклеотиди, докато не се достигне до триплета ЦЦА, а при други се прибавя тази последователност.
8.7. Синтеза на белтък (транслация или превеждане) на генетичната информация
Съхранението и предаването на генетичния материал са единствените познати функции на ДНК. Аминокиселинната последователност на всеки белтък в клетката, както и нуклеотидната последователност на всяка РНК се определят от нуклеотидната последователност на клетъчната ДНК. Сегмент ДНК, който съдържа информация за синтезата на функционален биологичен продукт (белтък или РНК) се нарича ген. Обикновено клетките съдържат хиляди гени и съответно ДНК е огромна молекула.
Информацията, записана в ДНК под формата на тройки нуклеотиди, се презаписва в иРНК. Матричната РНК (иРНК) е междинен преносител на генетичната информация от един или няколко гена до рибозомата, където се изгражда съответният белтък. В кибернетиката изобразяването на едни обекти чрез други се нарича кодиране. Тъй като триплетът „изобразява" мястото на една аминокиселина, той се нарича кодон (кодираща единица). Всеки презаписан кодон в иРНК се разпознава от свойствена специфична тРНК. Мястото в тРНК, което се разпознава от иРНК се състои също от три нуклеотида, които са комплементарни на нуклеотидите, образуващи кодона и се нарича антикодон.
Молекулата на ДНК е изградена от комбинирането на 4 нуклеотида, които се повтарят в различни комбинации. Аминокиселините в белтъчните молекули са 20. Ако допуснем, че последователността на нуклеотидите в молекулата на ДНК определя подреждането на аминокиселините в полипептидната верига, то не е възможно един нуклеотид да определя мястото на една аминокиселина. Ако това се осъществява от 2 последователни нуклеотида, възможни са 42=16 комбинации. Те също не са достатъчни. Франсис Крик доказа, че З последователни нуклеотида от веригата на ДНК, наречени триплет, определят мястото на 1 аминокиселина в полипептидната верига.
Мястото на всяка аминокиселина в полипептидната верига се определя от три последователни нуклеотида в молекулата на ДНК. Възможните комбинации от различни триплети са 43=64. От тях 61 определят мястото на 20-те протеиногенни аминокиселини, а 3 са безсмислени.
Генетичния (биологичен) код е начин за кодиранена информацията за строежа на белтъците във вид на нуклеотидна посредователност. Той е предназначен за да преведе четиризначния език на нуклеотидите (А, Г, У, Ц), на двадесетзначния език на аминокиселините. Той притежава някои особености:
- триплетност – три нуклеотида формират кодон, който кодира една аминокиселина. Общо има 61 смислени кодони.;
- специфичност (или еднозначност) – на всеки кодон съответства само една аминокиселина.;
- изроденост – за една аминокиселина може да има няколко кодона;
- универсалност – биологичния код е еднакъв за всички видове организми на Земята (само в митохондриите на млекопитаещите има изключение);
- колинеарност – последователността на кодоните съответства на последователноста на аминокиселините в кодирания белтък;
- непрепокриваемост – триплетите не се препокриват един с друг, а се разполагат последователно, един до друг;
- отсъствие на препинателни знаци – между триплетите няма допълнителни нуклеотиди или някакви други сигнали;
- еднопосочност – при синтезата на белтък, четенето на кодоните върви последователно, без пропуски или връщане назад.
Биологичният код не се проявява без съдействието на допълнителни молекули, които изпълняват свързваща функция.
Транспортните молекули РНК са единствените посредници между 4-ри буквената последователност на нуклеиновите киселини и 20-ет буквената последователност на белтъците. Всяка транспортна РНК има определена триплетна последователност в антикодоновата бримка (антикодон) и може да присъедини само една аминокиселина, която съответства на този антикодон. Именно от наличието един или друг антикодон в тРНК зависи, каква аминокиселина се включва в белтъчната молекула, т.е. нито рибозомата, нито иРНК разпознават аминокиселината. Функците на тРНК се изразяват в:
- специфично свързване с аминокиселините;
- специфично свързане с иРНК, съответно чрез взаимодействието кодон-антикодон;
- включване на аминокиселини в белтъчната верига в съответствие с информацията от иРНК.
Присъединяването на аминокиселините към тРНК се осъществява от ензим аминоацил-тРНК-синтетаза, който притежава специфичност към две съединения едновременно: към някоя аминокиселина и съответстващата на нея тРНК. Аминокиселината се присъединява към 3’ – края на акцепторната бримка на тРНК чрез своята α-карбоксилна група (фиг.122), връзката между аминокиселината и тРНК е макроергична. α-Аминогрупата остава свободна.
Съществуват около 60 различни тРНК, някои аминокиселини могат да се присъединяват към новосинтезиращата се полипептидна верига от две или три тРНК. Различни тРНК, които присъединяват една и съща аминокиселина се наричат изоакцепторни.
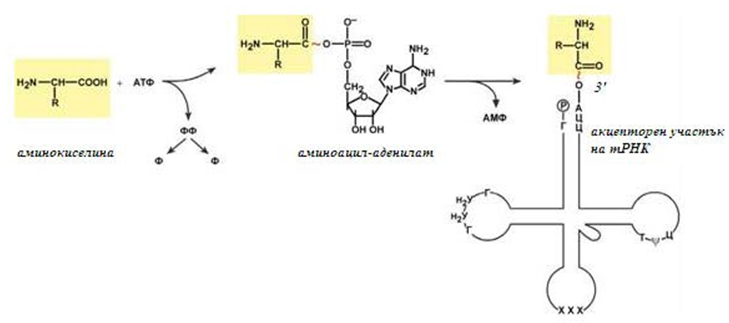
Фигура 122. Реакция на синтеза на аминоацил-тРНК
Транслацията е процес на биосинтез на белтък от матрицата на иРНК. След преноса на информацията от ДНК на матричната РНК, започва синтеза на белтък. Всяка зряла иРНК носи информация за ситезата само на една полипептидна верига. Ако на клетката са необходими други белтъци, то е необходима да се синтезират иРНК от други участъци на ДНК.
Биосинтезата на белтъци или транслацията протича в рибозомите, вътреклетъчни белтъксинтезиращи органели и включва:
- матрица – матрична РНК;
- растяща верига – полипептид;
- субстрат за синтез – 20 протеиногенни аминокиселини;
- източник на енергия – ГТФ;
- рибозомни белтъци рРНК и белтъчни фактори.
Синтезата се разделя на три основни етапа – инициация, елонгация и терминация.
Инициация. За инициацията са необходими иРНК, ГТФ, малка и голяма рибозомни субединици, три белтъчни фактора за инициация (ИФ-1, ИФ-2, ИФ-3), метионин и тРНК за метионина. В началото на този етап се формират два тройни комплекса:
- първи комплекс – иРНК, малка рибозомна субединица, ИФ-3;
- втори комплекс – метионил- тРНК, ИФ-2, ГТФ.
След формирането на тройните комплекси, те се обединяват с голямата суединица (фиг.123). В процеса активно участват белтъчните фактори за инициация, като източник на енергия служи ГТФ. След събирането в комплекс иницииращата метионил-тРНК се свързва с първия кодон АУГ на матричната РНК и се разполага в П-центъра (пептидилния център) на голямата субединица. А-центъра (аминоацилния център) остава свободен, той ще се задейства на следващия етап (елонгация), при свързването на аминоацил-тРНК. След присъединяването на голямата субединица започва етапа елонгация.
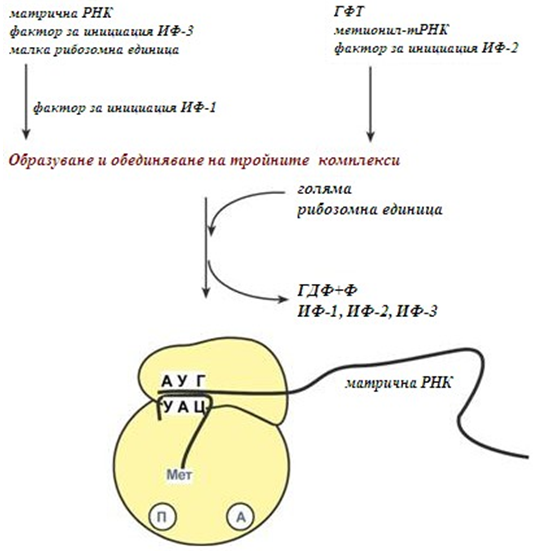
Фигура 123. Етап инициация - протичащи реакции
Елонгация. За този стадиий са необходими всичките 20 аминокиселини, тРНК за всички АК, белтъчните фактори за елонгация, ГТФ. Удължаването на веригата протича със скорост около 20 аминокиселини в секунда. Елонгацията представлява цикличен процес от три повтарящи се процеса (фиг.124):
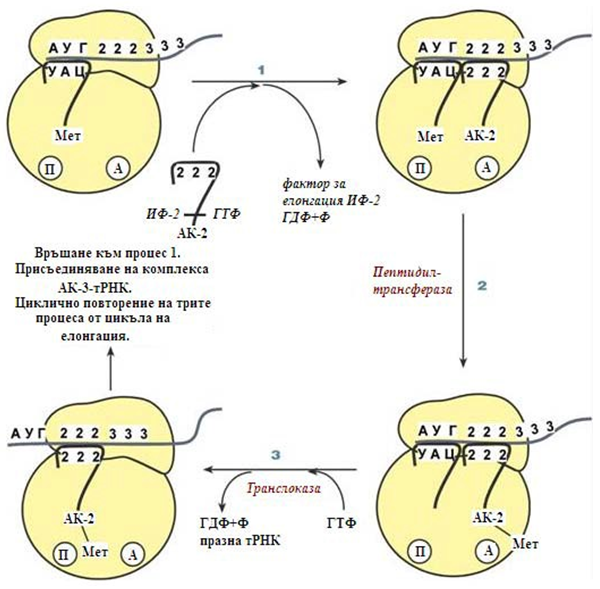
Фигура 124. Етап елонгация – циклично протичащи се реакции
- Присъединяване на аминоацил-тРНК към кодона на иРНК, АК се разполага в А-центъра на рибозомата, като източник на енергия служи ГТФ.
- Ензима пептидилтрансфераза осъществява пренос на метионина с метионил-тРНК (в П-центъра) на втората аминоацил-тРНК (в А- центъра) с образуване на пептидна връзка между метионина и втората АК. При това активираната СООН-група на метионина се свързва със свободната Nh3-група на втората АК, източник на енергия е макроергичната връзка между АК и тРНК.
- Ензима транслоказа премества иРНК относно рибозомата по такъв начин, че първия кодон АУГ се оказва извън рибозомата, втория кодон застава срещу П-центъра, а срещу А-центъра застава третия кодон. За този процес е необходима енергия, която се получава от ГТФ. Едновременно с преместването на иРНК се премества и закрепената на нея тРНК, така иницииращата първа тРНК излиза от рибозомата, а втората тРНК с дипептида се разполага в П-центъра.
Цикъла на елонгация (реакции 1, 2, 3 ), се повтаря до тогава, докато се синтезира полипептидната верига. Второто повторение на цикъла започва с присъединяване на третата аминоацил-тРНК към третия кодон на иРНК, АК-3 се помества в А-центъра, после трансферазната реакция се повтаря и се
образува трипептид, който заема А-центъра, след което се премества в П- центъра чрез транслоказна реакция. В освободения А-център влиза аминоацил-тРНК на четвъртата АК и започва третия цикъл на елонгация.
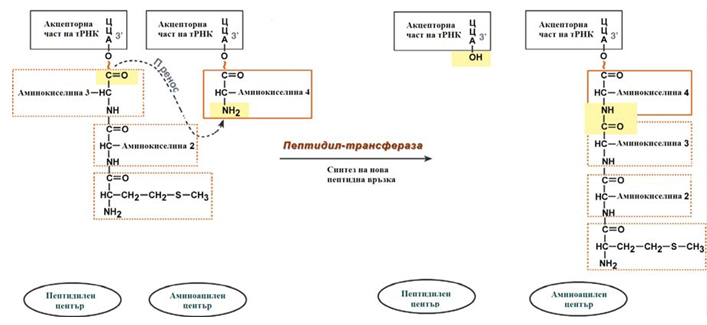
Фигура 125. Образуване на пептидна връзка в рибозомата
Терминация. Синтезата на белтък продължава, докато рибозомата не достигне на иРНК, до особен терминиращ кодон, или т.нар. стоп-кодон УАА, УАГ, УГА. Тези триплети не кодират нито една АК, наричат ги още нонсенс- кодони.
При влизане на тези триплети в рибозомата се активират белтъчни фактори за терминация, които последователно катализират:
- Хидролизисно отделяне на полипептида от последната тРНК.
- Отделяне на последната празна тРНК от П-центъра.
- Дисоциация на рибозомата. Източник на енергия е ГТФ.
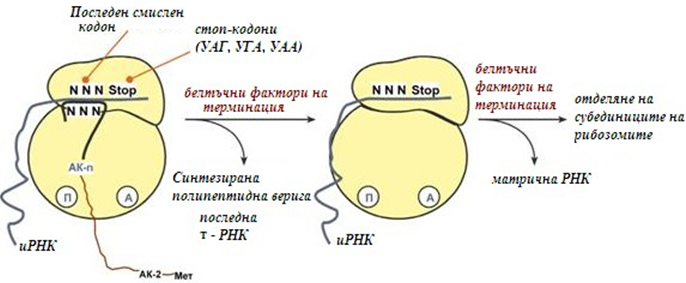
Фигура 126. Реакции на стадий терминация
Полирибозоми. За да се осъществи по-ефективен белтъчен синтез, към една иРНК понякога могат да се прикрепят няколко рибозоми една след друга, които синтезират еднакви копия от полипептидни вериги. Тези образувания се наричат полирибозоми.
8.8. Постъпване на нуклеотиди в организма
Нуклеотидите постъпват в организма, основно чрез храната в състава на нуклеопротеини. След въздействие на солната киселина и протеолитичните ензими в стомаха се разпадат до нуклеинови киселини и белтъчна част. Белтъците се храносмилат по свойствения за тях метаболитен път (фиг.127).
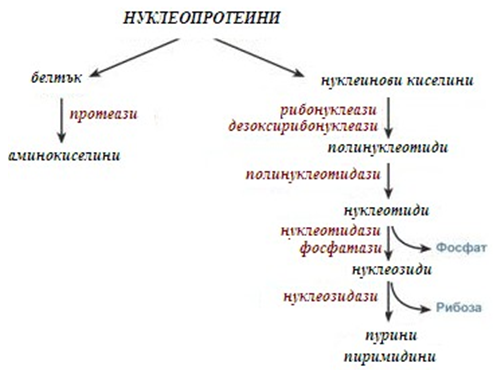
Фигура 127. Храносмилане на нуклеопротеините в стомашно-чревния тракт
Панкреатичният сок съдържа рибонуклеази и дезоксирибонуклеази, които хидролизират всички нуклеинови киселини до полинуклеотиди (фиг.127). След действието на панкреатичните ензими полинуклеотидазите (фосфодиестерази) от тънкото черво хидролизират нуклеиновите киселини до мононуклеотиди. Под действие на нуклеотидази и фосфатази протича хидролиз на нуклеотидите до нуклеозиди, които или се абсорбират, или под действието на нуклеозидази от чревната лигавица се разграждат до пурини и пиримидини.
В чревния лумен пуриновите бази могат да се окислят до пикочна киселина, която се всмуква и отделя с урината. По-голямата част от пурините, които се абсорбират от ентероцитите, също се превръщат в пикочна киселина, при това не протича включването им в образуващите се молекули на нуклеотиди и нуклеинови киселини. Свободните пиримидини, както и пурините, основно се катаболизират и отделят без да се използват от организма.
По такъв начин, нуклеиновите киселини от храната не постъпват от тънкото черво в кръвния поток и не се използват като непосредствени предшественици на ДНК и РНК от клетките на организма.
Към пуриновите азотни бази се отнасят аденин и гуанин. Хипоксантин, ксантин и пикочната киселина са крайни продукти от обмяната на пурините в организма на човек, повечето млекопитаещи и птиците.
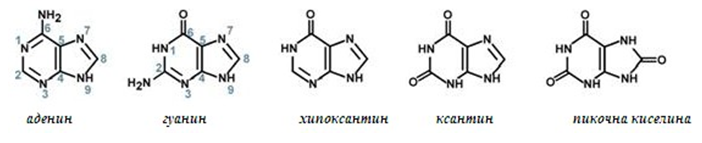
Фигура 128. Пуринови азотни бази и техните крайни продукти от катаболизма
Катаболизма на пуриновите бази може да се раздели условно на 5 етапа (фиг.129):
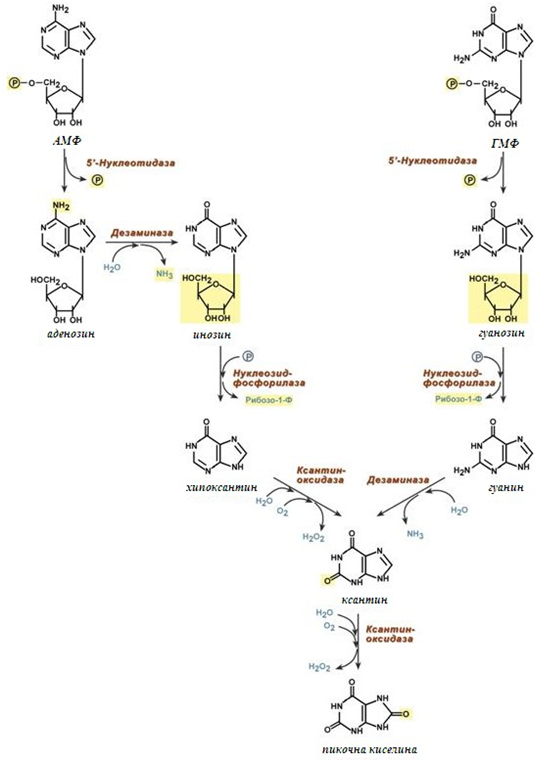
Фигура 129. Етапи на катаболизма на пуринови азотни бази
- Дефосфорилиране АМФ и ГМФ от ензима 5’-нуклеотидаза;
- Окисление на С6 в аденозина и едновременно дезаминиране от ензима дезаминаза, образува се инозин.
- Отделяне на рибозата от инозина (с образуване на хипоксантин) и гуанозина (с образуване на гуанин), с едновременно фосфорилиране от ензима нуклеозидфосфорилаза.
- Окисление на С2 в пуриновия пръстен, при това хипоксантин се окислява до ксантин (от ензима ксантиноксидаза), гуанин се дезаминира до ксантин от ензима дезаминаза.
- Окисление на С8 в ксантина с образуване на пикочна киселина от ензима ксантиноксидаза. Около 20% от пикочната киселина се отстранява от жлъчката в червния тракт, където се разгражда от микрофлората до СО2 и вода. Останалата част се отделя чрез бъбреците.
Разграждането на пиримидините води до образуване на амоняк, СО2 и вода.
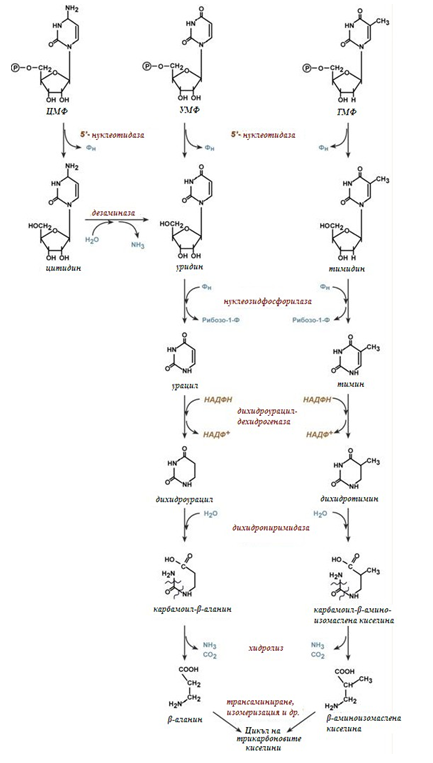
Фигура 130. Катаболизъм на пиримидиновите азотни бази
Ензима 5’-нуклеотидаза откъсва 5’-фосфатната група от ЦМФ, УМФ и ТМФ. Аденозин-дезаминаза осъществява окислително дезаминиране на цитидина. Нуклеозид-фосфорилаза отделя рибозата от уридина и тимина. Дихидроурацил-дехидрогеназата, възстановява урацила и тимидина. Дихидропиримидазата осъществява хидролиз на пиримидиновия пръстен. Протича хидролитично отделяне на амоняк и СО2. След окончателното разграждане на пиримидиновия пръстен, образуваните β-аминокиселини се подлагат на трансаминиране, след което кетокиселините изомеризират и се разграждат в ЦТК.
Терминологичен речник
- #репликация - синтез на ДНК, протича в строго определен период от живота на клетката.
- #ген - сегмент от ДНК, който съдържа информация за синтезата на функционален биологичен продукт (белтък или РНК).
- #триплет – три последователни нуклеотида от веригата на ДНК, които определят мястото на 1 аминокиселина в полипептидната верига.
- #триплетност – три нуклеотида формират кодон, който кодира една аминокиселина. Общо има 61 смислени кодони.
- #транслация - процес на биосинтез на белтък от матрицата на иРНК.
- #транскрипция - процес на презаписване на данните от ДНК на РНК е (англ.
- transcription – презаписване) – биосинтез на РНК от матрицата на ДНК.
- #рибозомна РНК (рРНК) – 80% от клетъчната РНК, изгражда клетъчните органели – рибозомите, където се извършва синтезата на белтъците.
- #принципа на комплементарността - срещу всеки нуклеотид, съдържащ определена азотна база, застава съответният му комплементарен нуклеотид.
- #иРНК – информационна РНК или матрична РНК, записаната в ДНК генетична информация под формата на тройки нуклеотиди, се презаписва в иРНК. иРНК е междинен преносител на генетичната информация от един или няколко гена до рибозомата, където се изгражда съответният белтък.
- #тРНК - около 15% от клетъчната РНК. Тази РНК има специфична структура на детелинов лист, от една страна свързват определени аминокиселини, а от друга се свързват и с иРНК. Названието “транспортна” идва от функцията, която тези РНК изпълняват – пренос на аминокиселините до рибозомите, където се намира иРНК.
- #матричен биосинтез или матричен принцип, означава че клетката синтезира полимерни молекули, такива като нуклеинови киселини и белтъци, на основата на шаблон – матрица.
- #антикодон - място в тРНК, което се разпознава от иРНК състои се също от три нуклеотида, които са комплементарни на нуклеотидите, образуващи кодона.

